One of the things I’ve noticed in the discussions and debates regarding autism is the regular use of 'impending doom' (cue ominous music) by advocates to make their case. This use is not limited to one particular ‘side’ of the autism debate – it can be seen at both ends of the viewpoint ‘spectrum’. And while I can see why some may feel that appeals to otherwise 'impending doom' (cue thunder) may strengthen their case, in the end I think it harms all of us. There are enough legitimate points to be made in autism discussions without having to unduly exaggerate or fabricate. And misinformation - especially misinformation that casts autistics in a damaging light – can outweigh the benefits that advocates are seeking to achieve.
One of the recent examples that comes to mind - courtesy of a post in Michelle Dawson’s blog - is the article Autism needs hope, written by Margaret Wente in the Globe and Mail on Nov 21, 2006.
From the article:
"My son is in a mainstream class. He goes to other kids' birthday parties," says Jean Lewis. She is the mother of an autistic 12-year-old and also a director of a B.C. parents' lobby group. Where would her son be without treatment? "He wouldn't be living at home," she says. "Without treatment, kids like this are usually institutionalized by the time they're adolescents. They're living in restraints, living in diapers. They have to have their teeth removed because they bite."
Um, excuse me?
The BC parents lobby group in question is F.E.A.T. of B.C. (F.E.A.T. stands for Families for Early Autism Treatment). A major goal of F.E.A.T. of BC, as per its official petition, is to "amend the Canada Health Act and corresponding Regulations to include IBI/ABA therapy for children with autism as a medically necessary treatment and require that all Provinces provide or fund this essential treatment for autism".
My daughter (3yrs 6mos old) is in an IBI program five mornings per week. While she was diagnosed as ‘autism, at the severe end of the spectrum’, and is approved and on the waiting list for IBI funding (currently running 'neck and neck' with university acceptance in terms of timing), we currently pay for her treatment privately. We pay a lot – a lot more than 50% of the figure quoted in the article. And for the moment, we feel that it is worth it. As such, groups like FEAT should theoretically be natural allies, or at least fellow travelers.
(BTW, for those who want to criticize us for using IBI, you may want to read here and here first to find out what we’re actually doing and believe. As an example, the Bear is not in IBI to teach her to pass for 'normal'. I’m sure that there is plenty of scope to criticize us for what we ARE doing without having to make things up that we DO NOT DO or support.)
But to me a central issue for lobbying groups and advocates is how they portray autistics. One might suggest that the quote above was an isolated statement. But from a story about ABA in 2005 referring to Ms Lewis’s son:
"Now in grade five in a regular classroom, the boy is a busy 11-year-old. He bikes, skis, swims and even plays clarinet in his school orchestra. He's also, "an active participant in family life," says Mr Lewis, who also serves as the President of the Autism Society of British Columbia. "Whatever we do, he's part of it."
But…
"But if his son's treatment was stopped, Mr Lewis is convinced the child would need to be institutionalized, possibly immediately. "If not now, then shortly," he says."
And from an Autism Fact Sheet on FEAT BC’s website:
"Without treatment, autism is a lifelong affliction that results in 90% of afflicted individuals placed in institutions and residential facilities, facing an unfulfilling and bleak existence for both the individual and family members."
Clearly the view being promoted is that autistics are dependent on ABA to avoid an 'unfulfilling and bleak' life of institutionalization (without teeth?). So much for Floortime, RDI, and other therapies, or implicit learning. (sarcasm) I guess THOSE parents should start saving for institutionalization now... (/sarcasm)
Ms Wente’s article also mentioned the ‘Autism Every Day’ video, including the ‘in another life, I’d love to go out for a bagel.' quote, and "For a glimpse of a day in the life of autism parents" it refers readers to a link to the video.
Personally, I support government funding (and not just limited to IBI - I favour a multi-disciplinary approach) for the support and education of autistic children, as well as required support for autistic adults. In this, organizations such as F.E.A.T. of B.C. should theoretically have my support. But I object to their methods, which clearly involve portraying autistics in a negative light through the dissemination of obviously incorrect information. It should not be necessary to denigrate my daughter to secure funding. Nor should appeals for funding be reduced to 'pay up now or I'll cost you more later'. What about the intrinsic right to an education and equality of opportunity?
As I wrote previously about the ‘Autism Every Day’ video:
"I quite like the idea of a video to explain our lives to others. But I'd prefer that it didn't make my child (by association) look like a ‘nightmare’ inflicted on us (and potentially on others when they come in contact with her). As a parent, autism is hard and painful at times to deal with, but there is also a joy that we can feel from our children, and one that our children can feel, even if we cannot always see it - and I think that much of the time I can see it, written on my daughter’s face. I don't want people to shy away from the Bear, to be afraid of the disruption that they fear she may cause, or to pity her, but rather, I want them to understand her and accept her.
Asking for acceptance does not mean that I'm not going to work like hell to fix her SI, immune, GI, etc. issues. I have no problem with autistic thought per se, but with autistic 'issues' - it is these that I'm working to help her overcome, and I believe there is a link between her medical symptoms and these ‘issues’. Asking for acceptance also does not mean that I won’t try through IBI or other teaching methods to teach or enable her to learn the skills she needs to make her way in the world. Instead, acceptance means recognizing her as a human being worthy of the same respect and rights as anyone else. Implying that she is a nightmare to deal with is not only untrue, but does nothing to further this goal."
In the same way, I believe that the message (as distinct from the goal of funding) propagated by FEAT of BC also does nothing to further the goal of acceptance of my daughter as a human being with full rights and worthy of respect.
I wrote to Ms Wente, pointing out my issues with her article, and closing with the following:
"While it is too late to change your article, might I suggest that you take a second look at this issue, as time permits? I (and probably most parents of autistic children) definitely appreciate anyone speaking on our children’s behalf, but what I’m asking is that you do so in a way that more accurately reflects our children’s reality – good and bad - and ultimately the reality of the autistic adults that they will become. I’m definitely not suggesting that you sugar coat the presentation. The reality is what it is, and should be presented rather than hidden away. But I would also ask that you recognize that our children are human beings, worthy of an accurate portrayal."
I will update this post with any reply, providing due consideration for Ms Wente’s privacy. In the meantime, if anyone else wants to contact her to offer their opinions, her e-mail - posted at the bottom of the autism article - is mwente@globeandmail.com. If you do so, please play nicely. "You can catch more flies with honey..."
In the meantime, I'll start saving for dentures for the Bear. After all, what is a bear without teeth?
Friday, December 15, 2006
Monday, November 20, 2006
The Sneaky Bear - Another Example of 'Theory of Mind'?
As I wrote in a previous post, I would suggest that the Bear is at least occasionally demonstrating Theory of Mind, despite some expert opinions that ToM in autistics is impaired, deficient, or non-existent (especially at such a young age). Recently we had another interesting (to me at least) potential example.
From Wikipedia, Theory of Mind is generally described as:
“a specific cognitive capacity: the ability to understand that others have beliefs, desires and intentions that are different from one's own.”
Further:
“This theory of mind covers two separate concepts:
1. Gaining the understanding that others also have minds, with different and separate beliefs, desires, mental states, and intentions
2. Being able to form operational hypotheses (theories), or mental models, with a degree of accuracy, as to what those beliefs, desires, mental states, and intentions are.“
Based on the the Sally-Ann test, 'normal' children below the age of four and most autistic children (of all ages) do not demonstrate this capability. (The link and also a comment by Camille on the previous ToM post discuss some of the issues with this test.)
A week or so ago, the Bear (who is 3 yrs, 5 mos old) was working with her IBI instructor, and her snack (chopped up pear chunks) was sitting in her bowl, nearby but out of immediate reach. At one point the instructor had to leave the room for a moment. While she was gone, the Bear reached over into the bowl to grab and eat some pear chunks, something that she would never have done while the instructor was present. What was more interesting though is that as the Bear realized that the instructor was returning, she quickly sat back and put her hands down in front of her with a 'butter wouldn't melt in her mouth' look, hiding the fact that she had been reaching into the bowl. It was the gestural equivalent of "I didn't do nuthin'!" To be clear, she didn't merely stop when she sensed that the instructor was returning, but acted in such a way to actually hide her previous actions.
Given the two conditions above, the implication is that:
a) the Bear understood (albeit incorrectly) that the instructor did not know that she had taken some of her snack (i.e. different and separate beliefs and intentions), and
b) that she formed an operational hypothesis that concealment of her actions was a possibility, i.e. that she had knowledge that her instructor might not have and that she might be able to maintain this state of differential knowledge.
As in the previous ToM post, I would suggest - if my interpretation of the Bear's actions is correct - that she is demonstrating that a child diagnostically labeled as ‘autism, at the severe end of the spectrum’ is capable of Theory of Mind, and at a young age. Again, maybe the 'experts' aren't totally correct on this one?
From Wikipedia, Theory of Mind is generally described as:
“a specific cognitive capacity: the ability to understand that others have beliefs, desires and intentions that are different from one's own.”
Further:
“This theory of mind covers two separate concepts:
1. Gaining the understanding that others also have minds, with different and separate beliefs, desires, mental states, and intentions
2. Being able to form operational hypotheses (theories), or mental models, with a degree of accuracy, as to what those beliefs, desires, mental states, and intentions are.“
Based on the the Sally-Ann test, 'normal' children below the age of four and most autistic children (of all ages) do not demonstrate this capability. (The link and also a comment by Camille on the previous ToM post discuss some of the issues with this test.)
A week or so ago, the Bear (who is 3 yrs, 5 mos old) was working with her IBI instructor, and her snack (chopped up pear chunks) was sitting in her bowl, nearby but out of immediate reach. At one point the instructor had to leave the room for a moment. While she was gone, the Bear reached over into the bowl to grab and eat some pear chunks, something that she would never have done while the instructor was present. What was more interesting though is that as the Bear realized that the instructor was returning, she quickly sat back and put her hands down in front of her with a 'butter wouldn't melt in her mouth' look, hiding the fact that she had been reaching into the bowl. It was the gestural equivalent of "I didn't do nuthin'!" To be clear, she didn't merely stop when she sensed that the instructor was returning, but acted in such a way to actually hide her previous actions.
Given the two conditions above, the implication is that:
a) the Bear understood (albeit incorrectly) that the instructor did not know that she had taken some of her snack (i.e. different and separate beliefs and intentions), and
b) that she formed an operational hypothesis that concealment of her actions was a possibility, i.e. that she had knowledge that her instructor might not have and that she might be able to maintain this state of differential knowledge.
As in the previous ToM post, I would suggest - if my interpretation of the Bear's actions is correct - that she is demonstrating that a child diagnostically labeled as ‘autism, at the severe end of the spectrum’ is capable of Theory of Mind, and at a young age. Again, maybe the 'experts' aren't totally correct on this one?
Tuesday, November 14, 2006
The Bear Goes Trick or Treating
This was the first Halloween for the Bear. We dressed her as a Hawaiian hula girl, complete with a lei (double wrapped) around her neck, another lei around her waist, a grass skirt, and a garland of flowers on her ankle. Given that this is Canada, the outdoor version also included leotards and a heavy top (both very non-Hawaiian), plus a coat to hide it all. We would have also liked to put a garland on top of her head, but it would have had the same life expectancy as any hat that is not tied down.
The Bear’s IBI program put on a Halloween event for the kids, providing loot bags, and practicing trick or treating. The Bear is GFCF, so I made the ultimate sacrifice, and afterwards replaced the treats with fruit. The Bear seemed to be quite accepting of her costume, and was very interested in the fact that everyone else was wearing one too.
Momma Bear and I talked about whether we should take her out trick or treating. We live in an area that is not exactly urban. The nearest driveway is nearly 300m away from the foot of our driveway, and that doesn’t include the distance to walk up to each house and back (often a 100m or more). Unless we wanted to drive to an urban area, any trick or treating would have to be via a car. Instead, Momma Bear thought that Halloween would be a good opportunity to introduce the Bear to the X’s.
The X’s are a couple (with older children, I believe in University) who live across the street and a couple of properties down. Mr X was in politics for many years. I won’t say any more since it will a) identify the X’s (I’d like to respect their privacy) and b) more closely identify us. We often see them riding their horses along the street and wave to them, and they say hi when they see us out and about. Mr X drops a bottle of wine off every Christmas on behalf of the local riding club. We have also been to their house for a neighbourhood gathering, plus to see the start of a one of the local hunts. But they had never met the Bear. Given that she is part of the neighbourhood (and also a potential flight risk), Momma Bear thought we should let them meet her so that they would know who she was.
So we dressed the Bear up and I drove her (with instructions to just stop by quickly, say hi, and come back) down the street and up their driveway. I walked the Bear to the door and knocked. Mrs X and the family dog came to the door, and she was surprised to see a trick or treater. Given the neighbourhood distances, we were probably the first to come by in a decade. I reminded her who we were (never taking it for granted that anyone who doesn’t see me often will remember exactly who I am), and said I just dropped by to quickly introduce my daughter to them.
Mrs X said hi to the Bear, complimented her on her costume, and seemed to immediately take to her. I mentioned at that point (since the Bear wasn’t saying anything) that she did not yet speak. This interested Mrs X. She had recently finished writing a book with a character that was ‘selectively mute’ and had done some research on this subject. I mentioned that the Bear was autistic, and she said that she had done some reading about autism too, and hinted that the character was on the spectrum.
Mrs X said that she would go and get a treat for the Bear, at which point I said that there was no need. She was on a selective diet, and I had just brought her over to introduce her, so that when they saw her they’d know who she was. Mr X was not home to meet the Bear, but Mrs X really seemed to take a shine to her. She wouldn’t take no for an answer on the treat, and asked what she could give her. After listing a few things (no, no, no...) she suggested a box of raisins, which I said would be okay. She invited us inside while she went to get them. Her voice seemed to be searching for something else, and then she asked if the Bear liked rocking horses? I said I thought so, so she said that they had one and she would pull it out.
Meanwhile, the Bear decided to go walking. Before I could grab her, she walked off into the darkened house. Crap. I’m inside the doorway in my shoes, the Bear has just walked off into their house, and Mrs X is down the hall off the kitchen trying to pull out the rocking horse. Something had to give, so I apologized and walked in. Meanwhile, the Bear had circled the front hallway and came into the kitchen from the opposite side. I came up the other side, helped Mrs X pull out a large fur and wood rocking horse, and finally everyone was in the same place, including the dog, (who also took a friendly interest in the Bear).
We put the Bear on the horse, and she seemed to enjoy it. I wish that I had a camera with me, since the opportunity to get a picture of my daughter in Hawaiian winter dress on a rocking horse does not come along that often. I mentioned that she was quite the daredevil and liked amusement rides, going on them at the local park where we had season’s passes.
Mrs X appeared to be quite interested in the Bear, and seemed to want to do more. “Does she like horses?” I’m not really sure, I answered. “Well, I have to feed and water the horses later anyway, so would she like to see the barn?” Er, okay? I said that we’d only popped by to say hi and introduce the Bear, and didn’t want to be any trouble, but Mrs X ducked the opportunity to politely shoo us off. So off we went to the barn. I carried the Bear as the path was a bit mucky, and continued to carry her though the barn and stables.
The barn was quite large, with about 20 horses (both theirs and boarders). Mrs X took us round and introduced each horse and its history, one by one, including a mother with a new colt. The Bear did not smile much, but she was clearly interested. She watched and took everything in, and was quite content to be carried around the barn. She patted a couple of the horses (hand over hand) and bopped a couple of horses that appeared too interested in tasting her lei and her grass skirt. Mrs X remarked on several occasions that she looked quite observant and also quite intelligent (which is a good way to make her Dad feel proud). Mrs X knew someone else in the neighbourhood (largely defined) who had an autistic son, and talked about him and his experiences.
We had a good chat about a lot of things, including but not limited to the Bear, got a good tour of the barn and the indoor riding area, looked (unsuccessfully) for the barn cat, and watched Mrs X give hay and water to each of the horses. Again, she seemed quite taken with the Bear, and ducked the opportunities I gave her to politely usher us on our way.
After the tour, she walked us back to our vehicle, and said that we could come by any time. I mentioned that my wife and I were not horse people (we’re probably the only ones in the neighbourhood who are not, although Momma Bear has ridden before), so that this was all new to us. Mrs X then mentioned teaching the Bear to ride, and I’m pretty sure that she actually offered to teach her. I’m not sure if I heard that correctly, and I’m not going to push the point. Mrs X did also mention that riding was probably good for autistic children. Living in horse country at least gives the Bear more of an opportunity, but on that we will have to see if she is interested.
Anyway, to make a long story short, Mrs X was the perfect host, and we really appreciated the time and the kindness that she provided. After I thanked her and drove off, I realized that we had been there for an hour and a half. All in all it was a good Halloween. The Bear had a nice experience, and made a new friend.
We need more Halloweens.

(FYI, a) the picture is in mid-swing (I'm quite impressed with the camera), b) the slant on the background is real - it is a hill behind her, and c) given Canadian weather this time of year, this is a reasonably realistic representation of what her costume looked like after coats, etc. were added)
The Bear’s IBI program put on a Halloween event for the kids, providing loot bags, and practicing trick or treating. The Bear is GFCF, so I made the ultimate sacrifice, and afterwards replaced the treats with fruit. The Bear seemed to be quite accepting of her costume, and was very interested in the fact that everyone else was wearing one too.
Momma Bear and I talked about whether we should take her out trick or treating. We live in an area that is not exactly urban. The nearest driveway is nearly 300m away from the foot of our driveway, and that doesn’t include the distance to walk up to each house and back (often a 100m or more). Unless we wanted to drive to an urban area, any trick or treating would have to be via a car. Instead, Momma Bear thought that Halloween would be a good opportunity to introduce the Bear to the X’s.
The X’s are a couple (with older children, I believe in University) who live across the street and a couple of properties down. Mr X was in politics for many years. I won’t say any more since it will a) identify the X’s (I’d like to respect their privacy) and b) more closely identify us. We often see them riding their horses along the street and wave to them, and they say hi when they see us out and about. Mr X drops a bottle of wine off every Christmas on behalf of the local riding club. We have also been to their house for a neighbourhood gathering, plus to see the start of a one of the local hunts. But they had never met the Bear. Given that she is part of the neighbourhood (and also a potential flight risk), Momma Bear thought we should let them meet her so that they would know who she was.
So we dressed the Bear up and I drove her (with instructions to just stop by quickly, say hi, and come back) down the street and up their driveway. I walked the Bear to the door and knocked. Mrs X and the family dog came to the door, and she was surprised to see a trick or treater. Given the neighbourhood distances, we were probably the first to come by in a decade. I reminded her who we were (never taking it for granted that anyone who doesn’t see me often will remember exactly who I am), and said I just dropped by to quickly introduce my daughter to them.
Mrs X said hi to the Bear, complimented her on her costume, and seemed to immediately take to her. I mentioned at that point (since the Bear wasn’t saying anything) that she did not yet speak. This interested Mrs X. She had recently finished writing a book with a character that was ‘selectively mute’ and had done some research on this subject. I mentioned that the Bear was autistic, and she said that she had done some reading about autism too, and hinted that the character was on the spectrum.
Mrs X said that she would go and get a treat for the Bear, at which point I said that there was no need. She was on a selective diet, and I had just brought her over to introduce her, so that when they saw her they’d know who she was. Mr X was not home to meet the Bear, but Mrs X really seemed to take a shine to her. She wouldn’t take no for an answer on the treat, and asked what she could give her. After listing a few things (no, no, no...) she suggested a box of raisins, which I said would be okay. She invited us inside while she went to get them. Her voice seemed to be searching for something else, and then she asked if the Bear liked rocking horses? I said I thought so, so she said that they had one and she would pull it out.
Meanwhile, the Bear decided to go walking. Before I could grab her, she walked off into the darkened house. Crap. I’m inside the doorway in my shoes, the Bear has just walked off into their house, and Mrs X is down the hall off the kitchen trying to pull out the rocking horse. Something had to give, so I apologized and walked in. Meanwhile, the Bear had circled the front hallway and came into the kitchen from the opposite side. I came up the other side, helped Mrs X pull out a large fur and wood rocking horse, and finally everyone was in the same place, including the dog, (who also took a friendly interest in the Bear).
We put the Bear on the horse, and she seemed to enjoy it. I wish that I had a camera with me, since the opportunity to get a picture of my daughter in Hawaiian winter dress on a rocking horse does not come along that often. I mentioned that she was quite the daredevil and liked amusement rides, going on them at the local park where we had season’s passes.
Mrs X appeared to be quite interested in the Bear, and seemed to want to do more. “Does she like horses?” I’m not really sure, I answered. “Well, I have to feed and water the horses later anyway, so would she like to see the barn?” Er, okay? I said that we’d only popped by to say hi and introduce the Bear, and didn’t want to be any trouble, but Mrs X ducked the opportunity to politely shoo us off. So off we went to the barn. I carried the Bear as the path was a bit mucky, and continued to carry her though the barn and stables.
The barn was quite large, with about 20 horses (both theirs and boarders). Mrs X took us round and introduced each horse and its history, one by one, including a mother with a new colt. The Bear did not smile much, but she was clearly interested. She watched and took everything in, and was quite content to be carried around the barn. She patted a couple of the horses (hand over hand) and bopped a couple of horses that appeared too interested in tasting her lei and her grass skirt. Mrs X remarked on several occasions that she looked quite observant and also quite intelligent (which is a good way to make her Dad feel proud). Mrs X knew someone else in the neighbourhood (largely defined) who had an autistic son, and talked about him and his experiences.
We had a good chat about a lot of things, including but not limited to the Bear, got a good tour of the barn and the indoor riding area, looked (unsuccessfully) for the barn cat, and watched Mrs X give hay and water to each of the horses. Again, she seemed quite taken with the Bear, and ducked the opportunities I gave her to politely usher us on our way.
After the tour, she walked us back to our vehicle, and said that we could come by any time. I mentioned that my wife and I were not horse people (we’re probably the only ones in the neighbourhood who are not, although Momma Bear has ridden before), so that this was all new to us. Mrs X then mentioned teaching the Bear to ride, and I’m pretty sure that she actually offered to teach her. I’m not sure if I heard that correctly, and I’m not going to push the point. Mrs X did also mention that riding was probably good for autistic children. Living in horse country at least gives the Bear more of an opportunity, but on that we will have to see if she is interested.
Anyway, to make a long story short, Mrs X was the perfect host, and we really appreciated the time and the kindness that she provided. After I thanked her and drove off, I realized that we had been there for an hour and a half. All in all it was a good Halloween. The Bear had a nice experience, and made a new friend.
We need more Halloweens.

(FYI, a) the picture is in mid-swing (I'm quite impressed with the camera), b) the slant on the background is real - it is a hill behind her, and c) given Canadian weather this time of year, this is a reasonably realistic representation of what her costume looked like after coats, etc. were added)
Friday, November 03, 2006
Respect Meme
Jonathan and María Luján started a ‘Respect Meme’ a couple of weeks ago, and NotMercury joined in. Their versions are here, here, and here respectively. I thought that I would join the stampede and give it a go.
1. What is respect for others?
I quite like and agree with NotMercury’s answer to this:
"Respect for others comes when we recognize our differences and make an effort to treat others the way we wish to be treated in spite of our differences"
I also like María Luján’s point about empathy being the root of a true understanding. A lot of people confuse empathy with agreement, but agreement is not required. To me, empathy is an understanding – or at least a reasonable attempt at understanding – of the position of another. Understanding another’s views does not require agreeing with them, or even liking them. But it does require listening rather than ‘selective listening’ or prejudging.
There will undoubtedly be disagreements, and it is legitimate to raise and discuss them. But disagreements over ideas, concepts, and even 'facts' and their interpretations (some forget that even 'facts' can be open to interpretation, especially when it comes to their meaning, importance, and implications) should not be treated as an excuse to attack the person with whom one disagrees. Even in the case of disagreement over conduct, one should still attempt to confine oneself to commenting on the behaviour, not the person. People make mistakes, and an error in judgment or action does not automatically render one a person unworthy of respect.
Having said that, I do not think that everyone is deserving of respect. I would suggest that one should always give the other person the benefit of the doubt, i.e. treat people as deserving of respect until they clearly demonstrate otherwise. But some people do clearly demonstrate otherwise. And then one has to choose how to respond. More on this in Question 4.
2. What are things that appear to respect issues, but are not?
I agree with Jonathan that some cannot separate the argument from their personhood, and thus incorrectly see a challenge to their ideas as a personal challenge. Confining one’s challenge to the idea itself and not the person behind can help to minimize this, but this will still be an issue at times.
A related issue is that some ‘wrap themselves in the flag’. On one side, some autistics view a disagreement with their ideas as a challenge or an attack on all of those who are autistic. "If you disagree with me then you don’t respect autistics". The mirror image on the 'all autism=mercury poisoning' side is "If you don’t agree with me then you are abusing children". Er, no, on both counts.
Another point I would agree with is that of NotMercury, who answered this with "When intentions are offered as justification for irresponsible actions." Good intentions do not automatically make one right or excuse negative actions, and should not shield those negative actions from comment or judgment. And legitimately questioning those actions can also be accomplished in such a way as to not disparage the intentions behind them (assuming that the intentions are good).
3. Is this relevant to the autism discussion and why?
Respect is very relevant to the autism discussion. I would suggest that from the point of view of the neurodiversity community, respect – and the perception (and in a lot of cases the reality) of the absence of respect - is one of the driving reasons behind their participation in the debate. I would suggest that the ‘autism = mercury’ parents also see the debate – in their case with the government and with many in the scientific community – as a respect issue, and also lack respect for the ND point of view as well as for many of its proponents. And for the majority of parents and caregivers, the respect issue revolves around government and support agencies not providing the amount of care and support that is required to improve the quality of life and accommodation of those touched by autism (i.e. not respecting what they see as the conditions required to respect the right to proper and adequate support).
I would suggest that the actions of many in response to the lack of respect that they feel they should be accorded are negatively affecting the wellbeing of all. But of course, "They started it!", so they will rail against and wait for the ‘other’ to fix the issues rather than seeking to build consensus around points on which they can agree. I would only half jokingly suggest that all involved need a course in ‘Interest Group Accommodation’. As a hint, more success can often be had by sitting at the table where the decisions are being made than by banging on the door outside. The goal is to get to the table, not to impotently make noise. Has anyone ever heard of the politics of entryism?
Some may argue that ‘rights’ are not items to be negotiated. But that is not what I am saying. Instead, I’ll use the analogy of the U.S. and the Soviet Union during the Cold War. Neither side was particularly accommodating of the other, nor were they willing to cede their rights where interests clashed. And at least in the Soviet case the expectation was not of long term co-existence but of an eventual ‘inevitable’ victory of communism and elimination of the other side (in its existing political form) as an opponent. Yet the two sides were still able to find common ground and agreement on more than a few issues that they shared in common, to mutual gain, while still remaining true to their long term goals.
4. What can we do to help resolve these issues?
Regardless of whether the other person deserves respect, one's own conduct is a separate issue. As María Luján quoted me on her blog:
"The real test of moral conduct is how one acts in the face of provocation. Someone else's bad behaviour towards me gives me the right to respond and to defend myself, but it does not give me carte blanche to respond in kind."
That is not an easy statement to live up to, and I would not even begin to suggest that I am successful in doing so in life. But one has to start somewhere. If my moral code goes out the window whenever someone challenges me, then what is it worth? To me, the correct response to clearly improper and sustained conduct is to point out the behavioural failings while remaining civil and true to one’s own code of civil conduct. Easier said than done. But if I bring myself down to the level of behaviour that I am objecting to then what have I gained?
To me, it all boils down to ‘Why are we participating in this *debate*?’ I am here to learn, possibly to contribute something back, and to act as a representative for my daughter until she is ready to take on this role for herself. I stand a better chance of accomplishing these goals if others are willing to engage in a dialogue. I am not here to make enemies - this is counterproductive. Some may be angered by my opposition to their viewpoints. That cannot be helped. But I can try my best to ensure that no one has as an excuse to shut down dialogue that I have personally and maliciously attacked them.
So, on to my point. Regardless of whether I respect someone, I owe it to them (usually), myself (definitely) and my daughter (definitely) to conduct myself in a civil manner, to remain true to my own ideals and code of conduct. I would suggest that – ultimately – this is the more powerful response. To be clear, I’m not suggesting that one should not defend oneself, or refrain from criticizing the arguments of others. Far from it, and I believe that I am quite vigorous in defending myself. But I am suggesting that there is a big difference between defending oneself and sinking to the level of one’s opponent.
5. How well do you think this will be accomplished?
On a personal level, I will try my best to be respectful of others, and at a minimum remain civil. At times I will fail (and should be called on this), but this is not an excuse to not try.
From an autism community standpoint, my expectations are low in the short term. It doesn't take long to find posts that have descended into slanging matches, or even posts that started at that level. Longer term, I think that answers provided by science, combined with the fact that the current generation of autistic children will one day become adults and influence the debate - probably in ways that will surprise all ‘sides’ - will be positive developments.
One of the keys to altering the tone of the debate is 'understanding' (again, this is distinct from 'agreement'), which is currently in short supply. I believe that as we learn more about autism we will find more items on which we can agree. But that will be a very long process.
As a final note, the above should not be construed as suggestive of anyone in particular. I would say that I have no issues with anyone who has commented on my blog to date (not just because they have commented here, but because we have ultimately been able to engage in a reasonable dialogue over time, even when we disagree). While I have had run-ins with some in the past, I have tried not to let it get personal, and I have found that one can (at least so far) eventually get to engage in a reasonable dialogue. I hope that others have found the same with me. As suggested on my ‘About Me’, we don't need to agree: I learn the most from those I don't agree with. And I would like to keep the discussion going.
1. What is respect for others?
I quite like and agree with NotMercury’s answer to this:
"Respect for others comes when we recognize our differences and make an effort to treat others the way we wish to be treated in spite of our differences"
I also like María Luján’s point about empathy being the root of a true understanding. A lot of people confuse empathy with agreement, but agreement is not required. To me, empathy is an understanding – or at least a reasonable attempt at understanding – of the position of another. Understanding another’s views does not require agreeing with them, or even liking them. But it does require listening rather than ‘selective listening’ or prejudging.
There will undoubtedly be disagreements, and it is legitimate to raise and discuss them. But disagreements over ideas, concepts, and even 'facts' and their interpretations (some forget that even 'facts' can be open to interpretation, especially when it comes to their meaning, importance, and implications) should not be treated as an excuse to attack the person with whom one disagrees. Even in the case of disagreement over conduct, one should still attempt to confine oneself to commenting on the behaviour, not the person. People make mistakes, and an error in judgment or action does not automatically render one a person unworthy of respect.
Having said that, I do not think that everyone is deserving of respect. I would suggest that one should always give the other person the benefit of the doubt, i.e. treat people as deserving of respect until they clearly demonstrate otherwise. But some people do clearly demonstrate otherwise. And then one has to choose how to respond. More on this in Question 4.
2. What are things that appear to respect issues, but are not?
I agree with Jonathan that some cannot separate the argument from their personhood, and thus incorrectly see a challenge to their ideas as a personal challenge. Confining one’s challenge to the idea itself and not the person behind can help to minimize this, but this will still be an issue at times.
A related issue is that some ‘wrap themselves in the flag’. On one side, some autistics view a disagreement with their ideas as a challenge or an attack on all of those who are autistic. "If you disagree with me then you don’t respect autistics". The mirror image on the 'all autism=mercury poisoning' side is "If you don’t agree with me then you are abusing children". Er, no, on both counts.
Another point I would agree with is that of NotMercury, who answered this with "When intentions are offered as justification for irresponsible actions." Good intentions do not automatically make one right or excuse negative actions, and should not shield those negative actions from comment or judgment. And legitimately questioning those actions can also be accomplished in such a way as to not disparage the intentions behind them (assuming that the intentions are good).
3. Is this relevant to the autism discussion and why?
Respect is very relevant to the autism discussion. I would suggest that from the point of view of the neurodiversity community, respect – and the perception (and in a lot of cases the reality) of the absence of respect - is one of the driving reasons behind their participation in the debate. I would suggest that the ‘autism = mercury’ parents also see the debate – in their case with the government and with many in the scientific community – as a respect issue, and also lack respect for the ND point of view as well as for many of its proponents. And for the majority of parents and caregivers, the respect issue revolves around government and support agencies not providing the amount of care and support that is required to improve the quality of life and accommodation of those touched by autism (i.e. not respecting what they see as the conditions required to respect the right to proper and adequate support).
I would suggest that the actions of many in response to the lack of respect that they feel they should be accorded are negatively affecting the wellbeing of all. But of course, "They started it!", so they will rail against and wait for the ‘other’ to fix the issues rather than seeking to build consensus around points on which they can agree. I would only half jokingly suggest that all involved need a course in ‘Interest Group Accommodation’. As a hint, more success can often be had by sitting at the table where the decisions are being made than by banging on the door outside. The goal is to get to the table, not to impotently make noise. Has anyone ever heard of the politics of entryism?
Some may argue that ‘rights’ are not items to be negotiated. But that is not what I am saying. Instead, I’ll use the analogy of the U.S. and the Soviet Union during the Cold War. Neither side was particularly accommodating of the other, nor were they willing to cede their rights where interests clashed. And at least in the Soviet case the expectation was not of long term co-existence but of an eventual ‘inevitable’ victory of communism and elimination of the other side (in its existing political form) as an opponent. Yet the two sides were still able to find common ground and agreement on more than a few issues that they shared in common, to mutual gain, while still remaining true to their long term goals.
4. What can we do to help resolve these issues?
Regardless of whether the other person deserves respect, one's own conduct is a separate issue. As María Luján quoted me on her blog:
"The real test of moral conduct is how one acts in the face of provocation. Someone else's bad behaviour towards me gives me the right to respond and to defend myself, but it does not give me carte blanche to respond in kind."
That is not an easy statement to live up to, and I would not even begin to suggest that I am successful in doing so in life. But one has to start somewhere. If my moral code goes out the window whenever someone challenges me, then what is it worth? To me, the correct response to clearly improper and sustained conduct is to point out the behavioural failings while remaining civil and true to one’s own code of civil conduct. Easier said than done. But if I bring myself down to the level of behaviour that I am objecting to then what have I gained?
To me, it all boils down to ‘Why are we participating in this *debate*?’ I am here to learn, possibly to contribute something back, and to act as a representative for my daughter until she is ready to take on this role for herself. I stand a better chance of accomplishing these goals if others are willing to engage in a dialogue. I am not here to make enemies - this is counterproductive. Some may be angered by my opposition to their viewpoints. That cannot be helped. But I can try my best to ensure that no one has as an excuse to shut down dialogue that I have personally and maliciously attacked them.
So, on to my point. Regardless of whether I respect someone, I owe it to them (usually), myself (definitely) and my daughter (definitely) to conduct myself in a civil manner, to remain true to my own ideals and code of conduct. I would suggest that – ultimately – this is the more powerful response. To be clear, I’m not suggesting that one should not defend oneself, or refrain from criticizing the arguments of others. Far from it, and I believe that I am quite vigorous in defending myself. But I am suggesting that there is a big difference between defending oneself and sinking to the level of one’s opponent.
5. How well do you think this will be accomplished?
On a personal level, I will try my best to be respectful of others, and at a minimum remain civil. At times I will fail (and should be called on this), but this is not an excuse to not try.
From an autism community standpoint, my expectations are low in the short term. It doesn't take long to find posts that have descended into slanging matches, or even posts that started at that level. Longer term, I think that answers provided by science, combined with the fact that the current generation of autistic children will one day become adults and influence the debate - probably in ways that will surprise all ‘sides’ - will be positive developments.
One of the keys to altering the tone of the debate is 'understanding' (again, this is distinct from 'agreement'), which is currently in short supply. I believe that as we learn more about autism we will find more items on which we can agree. But that will be a very long process.
As a final note, the above should not be construed as suggestive of anyone in particular. I would say that I have no issues with anyone who has commented on my blog to date (not just because they have commented here, but because we have ultimately been able to engage in a reasonable dialogue over time, even when we disagree). While I have had run-ins with some in the past, I have tried not to let it get personal, and I have found that one can (at least so far) eventually get to engage in a reasonable dialogue. I hope that others have found the same with me. As suggested on my ‘About Me’, we don't need to agree: I learn the most from those I don't agree with. And I would like to keep the discussion going.
Friday, October 13, 2006
Autism and the Evolution of the Brain
In an earlier post I approached the subject of Autism, Genetics, and Evolution from the perspective of the requirements imposed by alleles and the theory of evolution. This post is an attempt to approach the subject from a different perspective, i.e. the evolution of the brain. My last post discussed autism and minicolumns, suggesting that a) ASD has a minicolumnar underpinning, b) this underpinning is required (i.e. no narrow minicolumns means no ASD), c) it originates in the first 40 days of fetal development (i.e. it is not itself acquired post-natally), d) that this difference falls within the normal range (i.e. that having it does not ‘cause’ a diagnosis, although it may very well result in diversity of thought and cognition, i.e. neurodiversity), e) that something else is therefore required (with no significant speculation as to what that something else may be, other than to generically label it as a ‘second hit’), and f) that research needs to prove or exclude causality among the population of the vulnerable, i.e. proving that something does not cause ASD in those who are invulnerable does not prove that it does not cause ASD in those who are vulnerable. This post builds upon the minicolumn post.
Dr Casanova also provided me with a copy of a work in progress on brain evolution and minicolumns (Casanova - Big Brains Manuscript, in preparation for submission), which has some interesting implications for autism, and prompted me to do some further reading. The same caveat applies as in my last post: I’m attempting to post about some of his work and add what I consider to be some of the implications. As before, bear in mind that I am not a neuroscientist, and as such there is the definite possibility that I have misinterpreted some of the ideas and findings. As such, any errors are mine. In addition, any implications beyond those explicitly stated in Dr Casanova’s research papers should not be attributed to Dr Casanova unless specifically noted.
Brain Size and Structure
A logical starting point in evaluating brain evolution is encephalization, or brain size. Statistical models have been built that compare body vs. brain size across species, from which one can derive an ‘expected’ brain mass based on body size. The actual brain mass divided by the expected brain mass yields an encephalization quotient (EQ). A result higher than one indicates a larger than expected brain mass, while a result less than one indicates a smaller than expected brain mass.
EQ is important as it allows for comparisons of brain sizes across species by automatically adjusting for body size. Larger animals would be expected to have larger brains, and elephants and some whales do have larger brains than humans. But after adjusting for body size, humans have much larger brains than other species. Using an EQ measure based on encephalization of insectivorous primates, humans have an EQ of 28.8, and there is a large gap between modern humans and all non-human primates, including the great apes (our closest relatives). This large gap is filled by the increasing encephalization of prehistoric hominid ancestors.
Encephalization has not affected all parts of the brain equally. The neocortex has been the primary beneficiary of this trend:
"The enlargement of the brain is not proportional; that is, all parts do not develop at the same rate. The neocortex is by far the most progressive structure and therefore used to evaluate evolutionary progress" (Stephan, 1972, p 174)
And Dr Casanova writes (Casanova - Big Brains Manuscript):
"It is the areal expansion of the isocortex and its connections that is ultimately responsible for primate encephalization. Ultimately, the simple addition of minicolumns is sufficient to explain cortical expansion, increased gyrification, and the subsequent parcellation and re-optimization of functional neural networks."
As brain size increases, connectivity requirements also change. Increases in cognitive capabilities may not result simply from increasing brain size or adding minicolumns, but rather, may be a function of brain connectivity. A key requirement for the brain is to generate a network of connections allowing for stable information processing while minimizing conduction and wiring costs. Connectivity is ‘expensive’ in terms of energy required to build and maintain connections. Bigger brains require both more connections and the interconnection of distant locations. Longer connections can result in signal delay/attenuation, and increase the probability of something going wrong. Increasing physical distance also raises the costs of connectivity, as the amount of energy required to maintain connections over longer distances is significant.
The answer to this connectivity issue appears to be at least threefold: modularization, gyrification, and parcellation.
By arranging neurons into minicolumns and driving connectivity between minicolumns, connectivity costs are reduced. Further organization of minicolumns into macrocolumns and then parcellation into functionally differentiated cortical regions also reduces overall connectivity requirements. The net effect is to allow a large number of cells to be connected by fewer axons and in more short-range connections.
Gyrification, i.e. the ‘folding’ of the cortex, reduces the physical distance between different areas and therefore the length of interconnecting fibers. Think of a piece of paper with two dots on it. The absolute distance between the two dots decreases as the paper is folded and the two dimensional surface in effect becomes three dimensional. Only one third of the human cortex is exposed to the surface, with the rest being found within sulci. Increased gyrification also increases the ratio of short range versus long rang connections within the brain.
Encephalization also results in an increase in parcellation of the brain into functionally differentiated cortical regions. Connectivity does not scale absolutely with brain size. As Dr Casanova writes: "increasing numbers of minicolumns impose a connectivity constraint, as non-adjacent or near-neighboring minicolumns become isolated by physical distance. Increasing distance limits connectivity as the amount of energy required to generate and maintain long-distance connections is substantial." Parcellation reduces the need for long distance connectivity by concentrating functions within locations. As encephalization increases, so does the amount of white matter dedicated to local connections. The result is a brain that is "not as equally densely connected relative to a smaller brain."
Within primates the corpus callosum, which connects the right and left hemispheres, generally becomes relatively smaller as isocortex size increases. As Dr Casanova writes, "Interestingly, at a gross anatomical level, perhaps the most obvious manifestation of increased parcellation is that the two cerebral hemispheres tend to be less densely interconnected and more independent." Connections between more distant regions are still required, and these may be maintained through specialized neurons, e.g. spindle neurons. As encephalization increases parcellation and decreases and/or channels connectivity into communication between larger units, there appears to be a corresponding increase in the number of spindle neurons to maintain functional communications.
The Costs of Encephalization
The human brain is expensive to build and maintain from an energy and nutritional standpoint. Kleiber’s law expresses the relationship between body mass and body metabolic requirements, i.e. resting metabolic energy requirements (RMR), also known as basal metabolic energy requirements (BMR). The equation is:
RMR = 70 * (W^0.75)
where RMR is measured in kcal/day, and weight (w) is measured in kg.
Mammalian brain size also scales with body mass, according to the formula:
E = 1.77 * (W^0.76)
where E is brain mass in grams. The similarity in the two exponential scaling coefficients is significant, implying that brain size and RMR are related, and a further inference is that the size of an individual’s brain is closely linked to the amount of energy available to sustain it (Foley and Lee, 1991).
While the brain is about 2.5% of our body weight, it accounts for 22% of our resting metabolism (Leonard and Robertson 1992, p 186). This is in sharp contrast to anthropoid primates using approximately 8% of resting metabolism for the brain, and other mammals using 3-4%. In contrast though, total human resting metabolism does not differ significantly from that of other mammals. So the question is, where does the metabolic energy to grow and sustain the human brain come from?
The answer is the ‘Expensive Tissue Hypothesis’, posited by Aiello and Wheeler, 1995. Aiello and Wheeler analyzed the ‘expensive’ (in terms of metabolic energy) organs in the body - the heart, kidneys, liver and GI tract – noting that together with the brain they accounted for the major share of total body BMR. Comparing the expected vs. actual size of these organs for an average 65 kg human, they found that there were significant differences in actual vs. expected size for both the brain and the gut.
They wrote (Aiello and Wheeler 1995, p 203-205):
"Although the human heart and kidneys are both close to the size expected for a 65-kg primate, the mass of the splanchnic [abdominal/gut] organs is approximately 900g less than expected. Almost all of this shortfall is due to a reduction in the gastrointestinal tract, the total mass of which is only about 60% of that expected for a similar-sized primate. Therefore, the increase in mass of the human brain appears to be balanced by an almost identical reduction in the size of the gastrointestinal tract…
"Consequently, the energetic saving attributable to the reduction in the gastrointestinal tract is approximately the same as the additional cost of the larger brain. "
Since the heart, kidneys and liver cannot be significantly reduced in size, due to their critical functions, to keep BMR at the expected level the higher energy costs of encephalization must be met by the gut. The implication for humans is that there had to have been an increase in dietary quality – e.g. more easily digested food, and the liberation of more energy/nutrients per unit of expended digestive energy - to allow for both a smaller gut and the reallocation of energy to encephalization.
Further evidence for the link between diet and brain size comes from the recent (in evolutionary terms) decrease in brain size in humans. Ruff, Trinkaus, and Holliday (1997) found that the human EQ reached its peak approximately 90,000 years ago, and has since remained fairly constant. But, absolute brain size has decreased by 11% since 35,000 years ago, with most of this decrease (8%) coming in the last 10,000 years. EQ has remained relatively constant because of an equivalent decrease in body size during the same timeframe.
So, what happened? The most plausible interpretation is that EQ is a genetically governed trait, and should not have changed materially in the last 10,000 years. This period has also seen the greatest social/cultural progress in human history, suggesting that the change is not the result of evolutionary selective pressure. Instead, the most likely change has been one of a shortfall in meeting nutritional requirements, in the form of one or more limiting factors preventing the body and brain from achieving their maximum potential development. The most obvious change in this period was the introduction of agriculture (i.e. the agricultural revolution), accompanied by a large rise in grain consumption and a significant drop in animal consumption (from perhaps 50% of diet to 10% in some cases).
The issue is presumably not caloric. Instead, the most plausible current hypothesis is that the decrease in animal consumption resulted in a consequent shortfall in consumption of preformed long-chain fatty acids (Eaton and Eaton 1998). For optimal growth the brain is dependent on the fatty acids DHA (docosahexaenoic acid), DTA (docosatetraenoic acid), and AA (arachidonic acid), which constitute over 94% of all HUFA (highly unsaturated fatty acids) in human and mammalian gray matter (Eaton et al 1998). Neuronal membranes are composed of a thin double-layer of fatty acid molecules. Loss in DHA concentrations in brain cell membranes correlates to a decline in structural and functional integrity of this tissue (source).
There is some evidence that humans are not able to synthesize sufficient levels of EPA (eicosapentaenoic acid – another omega-3 fatty acid) or DHA from precursor alpha-linolenic acid (LNA), and thus must get these EFAs from their diet. An impaired ability to synthesize EPA or DHA would increase the dietary dependence. All of the above are far more plentiful in animal foods than plants. Analysis of likely EFA intake during prehistoric times under a wide range of assumptions suggests that levels of EFA would have been sufficient (0.9g/1000kcal) to allow brain expansion and evolution (Eaton et al 1998). As the human diet changed, our access to these essential fatty acids declined.
Eaton et al 1998 discuss the differences in Paleolithic vs. current dietary EFA consumption, comparing a composite ‘diet of evolutionary adaptedness’ or DEA to current EFA intakes. Prior to the agricultural revolution, human consumption of DHA is estimated at 270 mg/day, while AA consumption is estimated at 1810 mg/day. Current estimates for Western diets are 80 mg/day and 100-1000 mg/day respectively, and the discrepancy for DTA is assumed to be similar. Paleolithic C18 PUFA consumption is estimated at nearly 21.5 g/day, with LA (linoleic acid) at 8.84 g/day and LNA (linolenic acid) at 12.61 g/day, with an overall omega 6:3 ratio of 0.70 in the C18 category. Current Western consumption is LA at 22.5 g/day and LNA at 1.2 g/day, with a total omega 6:3 ratio of 16.74. The result is a massive variation from EFA consumption during the period of peak human brain size, and may result in a shortfall in availability of key EFAs. Why does this shortfall matter?
As Cordain et al, 2001 explains, all mammalian brain tissue appears to have an invariant requirement for DHA and AA, without which normal neural function cannot occur: "Limitations to the supply of either one of these fatty acids will determine limitations to brain growth." They also indicate that the supply of these EFAs is constrained by the limited ability of the liver as well as other tissue to synthesize these fatty acids from their dietary precursors (LNA and LA). As such, we must consume the EFAs we need to build brain tissue. As the authors state: "Encephalization quotients decrease with increasing body size because there literally may be insufficient long chain fatty acid product (AA and DHA) to build more brain tissue."
To add a further note, while the researchers above have concentrated on the implications of EFA changes in our diets, it is also reasonable to assume that the changes in diet with the agricultural revolution would also impact access to other key nutrients, minerals, and even essential and conditionally essential amino acids, as discussed here. Access to zinc, iron, taurine, natural Vitamin A (as distinct from beta carotene) and B12 are some of the nutrients that would have been significantly impacted by the change in diet, although these changes have not been explored to nearly the same extent as the EFA issue. This is not to comment on the implications of these changes or to suggest any consequences or causality, other than to note that they exist and may have implications beyond those discussed in this post.
The increased cost of encephalization has also had another consequence for human development – delayed maturity. As part of their analysis, Foley and Lee, 1991 compare the development of human and chimpanzee brain growth patterns. Chimpanzees are born with a brain mass that is 47% of adult brain mass, and adult size is reached by four years of age. Humans, in contrast are born with brain masses that are 25% of adult brain mass, and by four years old have reached only 84.1% of adult mass. Despite a slower maturation, human daily brain energy costs start at twice those of chimpanzees, and by age five are 3.3 times higher. Growth costs are also commensurately higher.
Foley and Lee suggested that “Mothers from a variety of primate (and other mammalian species) have a goal of weaning infants at an optimal mass, ensuring those infants’ survival, and themselves producing again.” They theorize that the reason for delayed maturity in humans was an inability to sustain both higher brain energy costs and high rates of brain growth, leading to evolutionary selection for the energetically less demanding strategy of increasing the duration of human growth and maturity. Despite a higher DQ and increased foraging efficiency (through increased socialization and cooperation over time), human brain energy costs were sufficiently high to force a slowdown in the rate of growth, despite the cost of reducing female lifetime reproductive rates. Larger brains forced slower growth rates and delayed human maturation.
Brain Evolution and Autism
The evolutionary factors described above have some implications for autism. First, as I stated in the prior post, analysis has determined that minicolumns in the brains of autistic individuals tend to be smaller in size, although with the same total number of cells per column (Casanova et al, 2002a and Casanova et al 2002b). Given that autistic brains tend to be larger than average, the results indicate that autistics also have a higher number of minicolumns. In addition, the neurons within these individual minicolumns tend to be reduced in size. Reduced minicolumn width appears to be a prerequisite for autism. But, the reported minicolumn widths found within autistic brains are still within the normal distribution of minicolumnar width, albeit at the tail end (Casanova 2006). For lack of a better term, I’ll refer to this as a ‘pre-autistic brain’, i.e. meeting the structural requirements for idiopathic autism but not qualifying for an ASD diagnosis (perhaps the broader autism phenotype or BAP?). I’ll also assume that the pre-autistic brain neurons are also of reduced size, for reasons that I will discuss below.
Studies have also shown a postnatal acceleration in brain growth in autistics, resulting in increased brain volume in autistic children vs. controls (e.g. Redcay and Courchesne, 2005), Aylward et al 2002, among others). As per Hazlett et al, 2006, "Significant enlargement was detected in cerebral cortical volumes but not cerebellar volumes in individuals with autism. Enlargement was present in both white and gray matter, and it was generalized throughout the cerebral cortex. Head circumference appears normal at birth, with a significantly increased rate of HC growth appearing to begin around 12 months of age. CONCLUSIONS: Generalized enlargement of gray and white matter cerebral volumes, but not cerebellar volumes, are present at 2 years of age in autism. Indirect evidence suggests that this increased rate of brain growth in autism may have its onset postnatally in the latter part of the first year of life." Thus the autistic brain appears to have more minicolumns, more neurons generally, and appears to grow faster than NT brains.
Given that encephalization appears to result from the addition of more minicolumns, with a resulting increase in gyrification and parcellation, could a more densely packed autistic or pre-autistic brain be a continuation of this evolutionary trend? If so, then the adaptations related to encephalization would also be expected to increase, e.g. increased gyrification and altered parcellation. This appears to be the case. Hardan et al 2004 reported an increase in left frontal gyrification in autistic children and adolescents compared with controls. Herbert et al 2004 reported differences in the parcellation of white matter (Casanova - Big Brains Manuscript), as well as increases in cerebral white matter (Herbert et al, 2003). More minicolumns require more connectivity, requiring more white matter, increasing total brain size, and increasing the bias toward local over global connectivity. The smaller neuronal size within the autistic brain would further reinforce this local connectivity bias.
It may also be noteworthy that the larger than average brains of autistic individuals have consistently shown smaller corpus callosi. Basically, a 'pre-autistic' brain is potentially a more cell-dense brain that may have advantages but is less robust. The addition of more minicolumns could explain the larger brain, increased gyrification, and some variations in parcellation, all within the normal range (albeit at the tail end of the distribution curve), and all of the above would help explain an increased preference of this brain for local over global processing (continuing the encephalization-linked trend), even without any further impact that would tip this still non-autistic brain into ASD (hence the term 'pre-autistic').
I would hypothesize that this pre-autistic brain is another variation along the path of increased encephalization, created by genetic lottery to either survive and reproduce - if this 'model' ultimately has an absolute advantage or niche advantage - or wither if it does not. Could more minicolumns - but of narrower width - and more neurons but of reduced size be a 'normal' tradeoff related in part to metabolism and limits in connectivity and skull size? Larger neurons would have higher metabolic requirements, so smaller neurons might be a response to a resource constraint (i.e. they might have been larger if the resources to grow and sustain them were available). If the same number of minicolumns and neurons were of 'normal' size then this might also unduly strain connectivity and metabolic demands (connectivity over increasing distance results in increased metabolic requirements) and skull size past any reasonable limits (e.g. an increase in minicolumn width from 44.3 um to 67.8 um as per Figure 1 below would be a 53% increase). In this light, the pre-autistic brain can be viewed as an evolutionary attempt to increase the size of the brain within existing resource constraints.
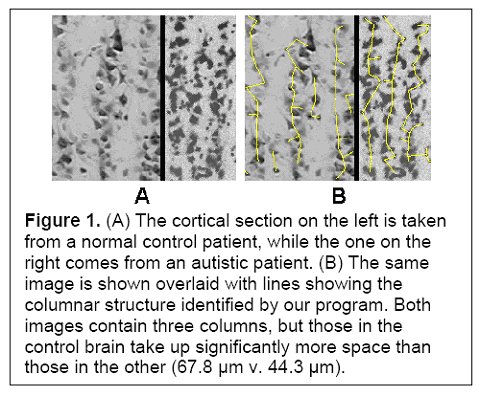
(Figure 1 Source – TMS Research proposal in preparation for submission, pg 2, with permission)
I would suggest that another reasonable hypothesis is that the pre-autistic brain could strain the nutritional capabilities of the body. If a normal brain constitutes 2.5% of body mass and accounts for 22% of BMR, (Wikipedia suggests that the developing infant brain consumes around 60% of the energy used by the body), then how much more would a larger and faster growing brain tax the metabolic system? Presumably a smaller neuron would require less energy or nutrition than a larger neuron, but I would suggest that the relationship is not linear, e.g. a neuron twice as large would require less than double the resources (in the case of the BMR, the exponential coefficient is 0.75, and it might be reasonable to expect something similar at a cellular level).
To the best of my knowledge it is an open question as to whether a pre-autistic brain also grows faster than average. A hypothesis is that the difference between the autistic and pre-autistic brain is the difference in growth rates. As stated in Courchesne, Redcay, and Kennedy 2004, “Head circumference, an accurate indicator of brain size in children, was reported to jump from normal or below normal size in the first postnatal months in autistic infants to the 84 th percentile by about 1 year of age; this abnormally accelerated growth was concluded by 2 years of age. Infants with extreme head (and therefore brain) growth fell into the severe end of the clinical spectrum and had more extreme neuroanatomical abnormalities.” Could increases in severity be a consequence of brain growth outstripping the required resources?
If autistic children have an increased rate of brain growth in the first few years of life then presumably this imposes a metabolic strain over and above the brain growth of NT children. Since no growth is ‘free’, this metabolic strain (energy and nutrients) must be somehow accounted for, either via an increased BMR, a reallocation of resources within the body, substitution (e.g. a deficiency in omega 3 EFA is compensated for by omega 6 EFAs, but at a functional cost), or through structural or operational deficiency. From Cordain et al, 2001 Fig. 2. “(Log-to-log chart of the resting metabolic rate of 20 anthropoids (new and old world monkey, apes and humans) relative to the predicted relationship based upon the Klieber equation)”, it is apparent that the actual to predicted BMR relationship is very close. I would suggest that BMR is a measure of efficiency that has been very tightly honed as a measure of evolutionary fitness (too low a BMR would result in energy constraints, while too high a BMR would increase the risk of starvation), and there is no reason to presume that autistics would vary significantly from non-autistics in adhering to expected results. But all of the other options result in potential downstream consequences in either the brain or other tissue.
If a shortfall in EFAs has potentially resulted in a decrease in size of the human brain by 11%, then what effect would the shortfall have on a pre-autistic brain with more minicolumns and an equivalent number of neurons per minicolumn – i.e. a brain with more neurons? Further, the increased rate of growth of the autistic (and pre-autistic?) brain in early childhood would increase even further the strain imposed by a shortage of EFAs. Smaller neurons, with a corresponding reduction in cell membranes and reduced metabolic demands might be a compensatory measure, gaining in metabolic efficiency and enhancing processing speed, the ability to process stimuli that require discrimination, and allowing for more complex information processing via local processing, but at a cost of reduced global processing and a reduction in ‘robustness’.
This tradeoff could still potentially have other consequences. Even with reduced neuronal size, the pre-autistic/autistic brain should presumably still require more energy and more nutrients than an NT brain, especially during the period of early childhood brain growth. If autistics adhere to the efficient tissue hypothesis then the logical tradeoff to satisfy this increased demand is with the GI system. And I would further speculate that another tradeoff may potentially occur with the liver.
Let’s be clear that I’m not suggesting at this point that the all autistic or pre-autistic children have GI or liver issues. What I am suggesting though is the following: a) that the increased brain costs must be met somehow, since no growth is ‘free’, b) that the GI system (and liver) are the logical organs to endure reductions to meet increased brain requirements, and c) that there in some cases may be consequences to this tradeoff in terms of the functional capabilities of these organs and potentially for overall health. In the case of the GI tract, some of these consequences could conceivably result in downstream issues (e.g. immune issues, impaired enzyme production, reduced metabolic efficiency, reduced nutrient and mineral absorption, etc.) that could have other systemic consequences, including consequences for the brain. Note that these issues would not ‘cause’ the brain to become ‘pre-autistic’, but they might have consequences for the pre-autistic brain that might result in an ASD diagnosis. Liver issues too could hypothetically have downstream consequences via a reduction in detox capabilities.
Further, if an NT brain is already facing resource constraints resulting in diminished size vs. potential (i.e. that 11% decrease in size), how much more critical would a GI 'issue' be for a pre-autistic brain? Even if the pre-autistic brain did not result in GI issues, comorbid GI issues might provide an additional resource constraint for a pre-autistic brain that could have a substantial impact on outcome. In this case it is not required that pre-autistic/autistic children have more GI issues than NT children, but rather, that given the increased metabolic demands of a pre-autistic/autistic brain, that the consequences of GI issues might be significantly greater on a larger and more demanding but less robust brain. The rates of comorbidity may be the same (although the efficient tissue hypothesis potentially suggests otherwise) but the consequences might not.
A further implication of the above goes back to the delayed maturity required of human offspring to allow for the management of the higher energy costs of encephalization. Since the autistic (and possibly the pre-autistic) brain is larger than an NT brain during the first few years of life and experiences a higher rate of growth, could delayed maturity be a further adaptive mechanism? Although much of this early growth is driven by connectivity, the increased number of neurons and minicolumns suggests that there is a lot more in the pre-autistic/autistic brain to connect, requiring substantial resources and energy. While this white matter growth is higher in autistics than NTs, it might still be a reduced rate of growth compared to what would be required to maintain an equivalent level of maturation in this larger brain as found in NT children.
Regardless of one’s beliefs regarding autism etiology, I would suggest that there are some serious implications regarding the nutritional status of autistics that flow from this analysis. The evolutionary and nutritional science referred to above is both mainstream and peer reviewed - no DAN! practitioners in sight. While the analysis has some similarities to Children With Starving Brains, the analysis, conclusions and hypotheses presented were arrived at from a different direction. I would suggest that it is a reasonable proposition that the autistic brain, if it is larger in childhood and goes through the growth spurt that mainstream research has detected, would have to face the same resource and energy constraints of NT brains, but with even higher requirements needing to be met. As such, one does not have to believe that addressing autistic nutritional requirements will result in a ‘cure’ to buy into the concept that autism brings with it some increased nutritional requirements, and that autistic brain ‘performance’ might be enhanced by compensatory nutritional intervention. Notice that I have ducked the entire issue of whether some autistic children with compromised diets due to sensory or tactile issues are further compounding any nutritional issues.
Finally, just to throw a hat into the discussion (see the last post’s comments regarding “The Meaning of Life” and hats for context), the above discussion may also offer a hypothesis to explain the ‘epidemic’, i.e. the perceived increase in the number of autistics in the past couple of decades. As mentioned above, absolute brain size has decreased by 11% since 35,000 years ago, with most of this decrease (8%) coming in the last 10,000 years. EQ has remained relatively constant because of an equivalent decrease in body size during the same timeframe. But in the post-WWII period human height has been increasing in the Western world and Asia. If EQ is genetically driven, then as stature increases, is brain size also on the rise? If resource constraints are lessening (perhaps with more variety in the diet, more access to protein, more access to EFAs?), then might constraints on pre-autistic brain size be diminishing along with constraints on human stature in general? Human stature can take at least two generations to recover (your grandmother’s dietary deficits affected your mother and the resources that she could bring to bear during your gestation). What if nutritional quality gains over the past two or more generations are now removing some of the nutritional constraints on pre-autistic brain development and size (i.e. more resources could allow for higher numbers of minicolumns to be generated during that first 40 days)? This might increase the pool of pre-autistic brains, i.e. the brains described above as being vulnerable to ASD.
Just something to think about.
Tuesday, September 05, 2006
Autism and Minicolumns
As a parent of an autistic child, a subject I find very interesting - and let’s distinguish up front between interest and comprehension or understanding - is ASD and brain research. To my mind, some of the more interesting work is being conducted by Dr. Manuel Casanova, at the University of Louisville, especially in the area of minicolumns. I have read some of Dr Casanova’s research and have written to him, and he has been gracious in replying and even forwarding other papers. I thought that I would make an attempt at a post about his findings and add what I consider to be some of the implications. Bear in mind that I am not a neuroscientist (no, really!) and as such there is the definite possibility that I have misinterpreted some of the ideas and findings. As such, any errors are mine. In addition, any implications beyond those explicitly stated in Dr Casanova’s research papers should not be attributed to Dr Casanova unless specifically noted.
I believe that autism has been ‘discovered’ in the wrong order. Autistic thought, loosely labeled as the broader autism phenotype (BAP), is a perfectly normal and reasonable example of neurodiversity, within the normal range of human thought. I see autistic disabilities as being driven by sensory integration issues to which those who fall within the BAP are more vulnerable. In this paradigm it is reasonable to expect the medical and psychiatric professions to ‘discover’ ASD in reverse order of severity, i.e. defining first the more severe disabilities, then lesser disabling cases, and then the population out of which these cases arise. It is also reasonable that the disabling ‘comorbidities’ of the BAP are therefore not essential features of this neurology, but rather, that they may be medical conditions worthy of amelioration, and that ameliorating these conditions would not change the underlying BAP way of thinking, but instead would allow it to flourish.
Minicolumns and The Brain
The neocortex (also known as the isocortex) is the 2 to 4mm tick top layer of the cerebral hemispheres and outer layer of the cerebral cortex. This layer is responsible for higher functions such as sensory perception, generation of motor commands, spatial reasoning, conscious thought, and language. The neocortex itself is divided into six layers, although there are no borders between these layers, and neuron dendrites and axons cross multiple layers.
The basic anatomical and physiological unit of the neocortex is the minicolumn. (Casanova 2006) As its name suggests, a minicolumn is a vertically aligned collection of cells and their projected connections. It is the smallest unit capable of information processing in the brain - resembling a mini-processor in function - receiving stimuli from elsewhere, processing information, and providing the capability of a response. Information is transmitted through the core of the minicolumn and is prevented from activating neighboring minicolumns by surrounding inhibitory fibers (interneuronal projections).
Pyramidal cells make up approximately 80% of the neurons of the cortex, and are the heart of the information processing capability of the minicolumn. Pyramidal cell somata (cell bodies) in layers III, V and VI are vertically oriented. Their dendrites and axon trees cross at least three layers, and in many cases all of the layers of the neocortex. They release glutamate as their neurotransmitter, and are the major excitatory component of the cortex.
Another component of minicolumns are various types of GABAergic inhibitory interneurons (i.e. neurons that produce GABA as their neurotransmitter), which tend to be aligned one on top of the other and modulate the activity of minicolumn pyramidal cells. Double bouquet cells are present in all layers, but are most dense in layers II and III. The axon bundles (long projections that conduct electrical impulses away from the cell body) of these cells are projected deep into the cortex from layer II to layer V, terminating on pyramidal cells as well as other inhibitory interneurons. They create a narrow vertical stream of inhibition through the cortex, as well as a vertically directed disinhibition of those pyramidal cells upon which the other inhibitory interneurons project. Other interneurons include Chandelier cells, which synapse directly onto the axon hillock (base of the axon projection from the cell body) of pyramidal cells, modulating cell output and participating in intra-columnar inhibition, and basket cells, which contacts the soma and dendrites of pyramidal cells, modulating input to these cells. Although these interneurons make up a small fraction of the total number of minicolumn cells, they play a significant role in finely tuning cortical information processing. A simplified representation of a minicolumn is thus of a processing core surrounded by a GABAergic interneuron circumferential zone of inhibitory and disinhibitory activity.
(Figure 2 Source – TMS Research Proposal in preparation for submission, pg 2, with permission)
(Figure 3 Source – TMS Research proposal in preparation for submission, pg 3, with permission)
Minicolumns are the basic organizational unit of the cortex. It is an increase in the number of minicolumns, expanding the area covered by the isocortex, that is responsible for the historical process of encephalization, i.e. the increase in size of the brain to a degree greater than that expected based on body size (in modern humans, brain size is three to five times greater than expected when comparing to mammals of equal body mass) (Casanova - Big Brains Manuscript, in preparation for submission). A variable number of minicolumns are dynamically clustered with neighbouring minicolumns into macrocolumns, and from there in networks of macrocolumns and regions of functional differentiation within the brain. The minicolumn-macrocolumn relationship may be linked in part to both the termination of projections from the thalamus, which span a fixed distance and may serve to link together minicolumns that receive input from the same thalamocortical fibers, and by the effects of serotonin, changing columnar development in the cortex during brain development (Casanova 2006).
This minicolumn organizational structure conveys certain advantages. Through limiting connectivity within the brain (connectivity requires a significant amount of energy), the result is a reduction in metabolic expenditure. Instead of connecting every cell within the cortex to different brain regions, projections are organized into modules. Single cells project to target sites, and information gained or transmitted is transferred to other neurons within the same modules. Another advantage is plasticity, or the ability of the brain to naturally reorganize connections. Similarities between minicolumns suggest that they are capable of performing similar transformations on incoming information, allowing them to replace each other in case of injury or to adapt to changing requirements. This is not to suggest that all minicolumns are identical. There are variations within different regions, but these may arise from variations in input, output targets, interconnectivity, and inhibition (Casanova - Big Brains Manuscript, in preparation for submission).
So, what does this have to do with autism?
Analysis has determined that minicolumns in the brains of autistic individuals tend to be smaller in size, although with the same total number of cells per column (Casanova et al, 2002a and Casanova et al 2002b). Given that autistic brains tend to be larger than average, the results indicate that autistics also have a higher number of minicolumns. In addition, the neurons within these individual minicolumns tend to be reduced in size.
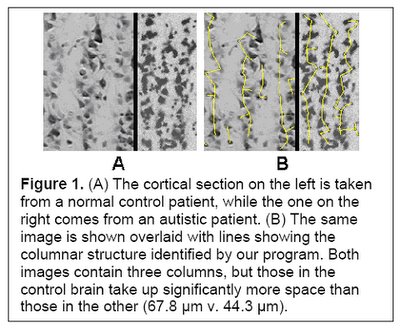
(Figure 1 Source – TMS Research proposal in preparation for submission, pg 2, with permission)
Smaller minicolumns would skew information processing (noise/signal) in favour of signal, potentially enhancing the ability to process stimuli that require discrimination, but also potentially at the expense of generalizing the salience of a particular stimulus. Smaller and more densely packed minicolumns could also allow for more complex information processing. As an example, the smaller minicolumns in the visual cortex may support added functionality, e.g. depth or color perception. This may be due to the overlap of neuronal projections allowed for when neuronal dendrites and axons remain the same size but the distance to neighbouring minicolumns is reduced (Casanova - Abnormalities Of Cortical Circuitry In The Brains Of Autistic Individuals).
These enhancements come at a cost. In autism, minicolumn size reductions result primarily from reductions in the peripheral zone of inhibitory and disinhibitory activity. The inhibitory fibers act to keep the stimuli within individual minicolumns, and the reduction in this space results in stimuli no longer being contained. Instead, they can overflow to adjacent minicolumns, providing an amplifier effect, which may explain hypersensitivity in some autistics. Thalamic input may also be amplified if thalamic terminal fields remain the same size and therefore result in more minicolumns per stimulated macrocolumn. Each minicolumn’s response to thalamic input is also modulated by the activity of neighbouring columns to a greater or lesser degree, allowing for gradations of response, so a reduction in GABAergic inhibitory activity could also result in a loss of inhibition and greater amplification (see Figure 1). Stimuli ‘spill’ and amplification could result in increased incidence of seizures in autistics.
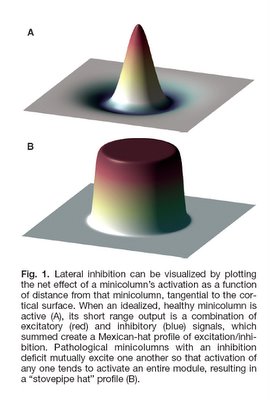
(Casanova 2006, pg 3, with permission)
An additional cost is that imposed by a reduction in neuron size. The ability of a neuron to sustain a connection over distance is related to the size of its cell body. Smaller neurons result in a metabolic bias favouring shorter connections at the expense of both longer distance and inter-hemispheral connectivity. The result is that autistic brains have a bias towards local (intra-regional) over global (inter-regional) connectivity and processing. Short intra-regional processing functions include mathematical calculations and visual processing. Cognitive functions that require inter-regional processing would be less metabolically efficient, including language, face recognition, and joint attention (Casanova - Abnormalities Of Cortical Circuitry In The Brains Of Autistic Individuals).
The total number of minicolumns in the human brain is defined during the first forty days of fetal development. Symmetrical divisions of germinal cells determine the total number of minicolumns, and a second phase of asymmetrical divisions provides for cell migration into the cortex, with successive divisions determining the number of cells in each minicolumn. This is a complex process, defined by a large number of genes interacting with the environment. The result is that a higher number of minicolumns is not something that someone can ‘acquire’ or be vulnerable to within the post-natal period. It is possible for disruptions during the narrow window of fetal development (e.g. rubella, thalidomide, tuberous sclerosis) to also result in an increase in the number of minicolumns, by influencing this process (Casanova - Abnormalities Of Cortical Circuitry In The Brains Of Autistic Individuals; Casanova 2006).
One point that is important to make is that while narrower minicolumns is a factor in autism, evidence suggests that it is not related to mental retardation. In Down Syndrome, as an example, minicolumn width is normal, despite the smaller brain sizes of Down Syndrome patients.
Implications
Reduced minicolumn width appears to be a prerequisite for autism. But, the reported minicolumn widths found within autistic brains are still within the normal distribution of minicolumnar width, albeit at the tail end (Casanova 2006). This suggests to me that the existence of narrow minicolumns is not enough by itself to result in an ASD diagnosis. The key instead appears to be a reduction in inhibition within minicolumns, rather than width alone. Reduced width increases the consequences of reduced inhibition, but does not automatically cause it. In effect, a brain with narrower minicolumns may be less robust, and therefore more vulnerable to the complications that could come with deviation from the narrow tolerances within which the brain functions. In a brain with wider minicolumns, a loss of inhibition would not have as significant an impact, as minicolumnar width (and therefore distance between minicolumn information processing cores) would still exist to reduce intercolumnal spill, and thalamic projections would result in fewer minicolumns per macrocolumn to be affected.
It is a logical assumption that minicolumn development in MZ twins would be similar (same alleles, very minimal epigenetic differences during the early weeks of fetal development). But MZ twins do not always share the same ASD diagnosis. In cases in which one twin has an ASD diagnosis, the chance of the second twin also having an ASD diagnosis (not necessarily of the same severity) is 60%, and the chance of the second twin being within the broader autism phenotype is 92%. While ASD clearly has an inheritable component (the prevalence of ASD in the population is 0.6%, but within families with an ASD child, the chance of subsequent children being ASD is up to 10%), genetics alone is not sufficient to explain a diagnosis in all cases. This, combined with the fact that reduced autism minicolumn widths still fall within the normal range suggests to me that the issue in autism relates to loss of inhibition, to which autistics are more vulnerable than average, i.e. reduced minicolumnar width is a precondition for autism but a 'second hit' is still required. If that second hit were purely genetic, as distinct from epigenetic or environmental, then presumably both MZ twins should be similarly affected. This is clearly not the case a significant percentage of the time.
Support for this may come from studies of the brains of MZ twins discordant for autism. In Kates et al 2003, a comparison of MZ twins discordant for autism indicated that both the autistic and non-autistic (broader autism phenotype) twins showed no significant differences in cerebral gray matter. This suggests a structural similarity in MZ twins, and the possibility of a requirement for a ‘second hit’ to explain the discordant outcomes.
We can speculate on what that ‘second hit’ may be, and indeed the speculation as to a) whether a second hit exists and b) the causes, if any, are among the most controversial subjects of debate within the community of those linked by autism. I will look a bit more closely in this direction in a post in the near future.
What is clear from above though is that if narrow minicolumnar width is a) a precondition for ASD and b) not sufficient by itself to cause ASD, then this should have an impact on ASD research. If narrow minicolumns creates a vulnerability that wider minicolumns do not, then demonstrating lack of effect in those who are invulnerable does not demonstrate lack of effect in those who may be vulnerable. In many cases the correct population in causation analyses (e.g. epidemiological studies?) would not be the wider population, but instead the sub-section of the population who are actually vulnerable. And in looking for causes of autistic disability, we may want to look more closely at those events and/or stimuli that may reduce inhibition within the brain and/or further discourage global connectivity.
It is also a reasonable proposition is that a ‘cure’ for autism would not alter the underlying narrow minicolumnar structure of the brain. Instead, it would allow this structure to function more effectively, maintaining its strengths while reducing or eliminating the more disabling features of ASD. A cure would not change who one is, which is one of the fears of the neurodiversity community. Instead, it would allow autistic thought to function without the disabling comorbidities, serving to maintain neurodiversity and allowing it to flourish.
Update - Related post:
Autism and the Evolution of the Brain (Oct 13, 2006)
I believe that autism has been ‘discovered’ in the wrong order. Autistic thought, loosely labeled as the broader autism phenotype (BAP), is a perfectly normal and reasonable example of neurodiversity, within the normal range of human thought. I see autistic disabilities as being driven by sensory integration issues to which those who fall within the BAP are more vulnerable. In this paradigm it is reasonable to expect the medical and psychiatric professions to ‘discover’ ASD in reverse order of severity, i.e. defining first the more severe disabilities, then lesser disabling cases, and then the population out of which these cases arise. It is also reasonable that the disabling ‘comorbidities’ of the BAP are therefore not essential features of this neurology, but rather, that they may be medical conditions worthy of amelioration, and that ameliorating these conditions would not change the underlying BAP way of thinking, but instead would allow it to flourish.
Minicolumns and The Brain
The neocortex (also known as the isocortex) is the 2 to 4mm tick top layer of the cerebral hemispheres and outer layer of the cerebral cortex. This layer is responsible for higher functions such as sensory perception, generation of motor commands, spatial reasoning, conscious thought, and language. The neocortex itself is divided into six layers, although there are no borders between these layers, and neuron dendrites and axons cross multiple layers.
The basic anatomical and physiological unit of the neocortex is the minicolumn. (Casanova 2006) As its name suggests, a minicolumn is a vertically aligned collection of cells and their projected connections. It is the smallest unit capable of information processing in the brain - resembling a mini-processor in function - receiving stimuli from elsewhere, processing information, and providing the capability of a response. Information is transmitted through the core of the minicolumn and is prevented from activating neighboring minicolumns by surrounding inhibitory fibers (interneuronal projections).
Pyramidal cells make up approximately 80% of the neurons of the cortex, and are the heart of the information processing capability of the minicolumn. Pyramidal cell somata (cell bodies) in layers III, V and VI are vertically oriented. Their dendrites and axon trees cross at least three layers, and in many cases all of the layers of the neocortex. They release glutamate as their neurotransmitter, and are the major excitatory component of the cortex.
Another component of minicolumns are various types of GABAergic inhibitory interneurons (i.e. neurons that produce GABA as their neurotransmitter), which tend to be aligned one on top of the other and modulate the activity of minicolumn pyramidal cells. Double bouquet cells are present in all layers, but are most dense in layers II and III. The axon bundles (long projections that conduct electrical impulses away from the cell body) of these cells are projected deep into the cortex from layer II to layer V, terminating on pyramidal cells as well as other inhibitory interneurons. They create a narrow vertical stream of inhibition through the cortex, as well as a vertically directed disinhibition of those pyramidal cells upon which the other inhibitory interneurons project. Other interneurons include Chandelier cells, which synapse directly onto the axon hillock (base of the axon projection from the cell body) of pyramidal cells, modulating cell output and participating in intra-columnar inhibition, and basket cells, which contacts the soma and dendrites of pyramidal cells, modulating input to these cells. Although these interneurons make up a small fraction of the total number of minicolumn cells, they play a significant role in finely tuning cortical information processing. A simplified representation of a minicolumn is thus of a processing core surrounded by a GABAergic interneuron circumferential zone of inhibitory and disinhibitory activity.

(Figure 2 Source – TMS Research Proposal in preparation for submission, pg 2, with permission)
(Figure 3 Source – TMS Research proposal in preparation for submission, pg 3, with permission)
Minicolumns are the basic organizational unit of the cortex. It is an increase in the number of minicolumns, expanding the area covered by the isocortex, that is responsible for the historical process of encephalization, i.e. the increase in size of the brain to a degree greater than that expected based on body size (in modern humans, brain size is three to five times greater than expected when comparing to mammals of equal body mass) (Casanova - Big Brains Manuscript, in preparation for submission). A variable number of minicolumns are dynamically clustered with neighbouring minicolumns into macrocolumns, and from there in networks of macrocolumns and regions of functional differentiation within the brain. The minicolumn-macrocolumn relationship may be linked in part to both the termination of projections from the thalamus, which span a fixed distance and may serve to link together minicolumns that receive input from the same thalamocortical fibers, and by the effects of serotonin, changing columnar development in the cortex during brain development (Casanova 2006).
This minicolumn organizational structure conveys certain advantages. Through limiting connectivity within the brain (connectivity requires a significant amount of energy), the result is a reduction in metabolic expenditure. Instead of connecting every cell within the cortex to different brain regions, projections are organized into modules. Single cells project to target sites, and information gained or transmitted is transferred to other neurons within the same modules. Another advantage is plasticity, or the ability of the brain to naturally reorganize connections. Similarities between minicolumns suggest that they are capable of performing similar transformations on incoming information, allowing them to replace each other in case of injury or to adapt to changing requirements. This is not to suggest that all minicolumns are identical. There are variations within different regions, but these may arise from variations in input, output targets, interconnectivity, and inhibition (Casanova - Big Brains Manuscript, in preparation for submission).
So, what does this have to do with autism?
Analysis has determined that minicolumns in the brains of autistic individuals tend to be smaller in size, although with the same total number of cells per column (Casanova et al, 2002a and Casanova et al 2002b). Given that autistic brains tend to be larger than average, the results indicate that autistics also have a higher number of minicolumns. In addition, the neurons within these individual minicolumns tend to be reduced in size.
(Figure 1 Source – TMS Research proposal in preparation for submission, pg 2, with permission)
Smaller minicolumns would skew information processing (noise/signal) in favour of signal, potentially enhancing the ability to process stimuli that require discrimination, but also potentially at the expense of generalizing the salience of a particular stimulus. Smaller and more densely packed minicolumns could also allow for more complex information processing. As an example, the smaller minicolumns in the visual cortex may support added functionality, e.g. depth or color perception. This may be due to the overlap of neuronal projections allowed for when neuronal dendrites and axons remain the same size but the distance to neighbouring minicolumns is reduced (Casanova - Abnormalities Of Cortical Circuitry In The Brains Of Autistic Individuals).
These enhancements come at a cost. In autism, minicolumn size reductions result primarily from reductions in the peripheral zone of inhibitory and disinhibitory activity. The inhibitory fibers act to keep the stimuli within individual minicolumns, and the reduction in this space results in stimuli no longer being contained. Instead, they can overflow to adjacent minicolumns, providing an amplifier effect, which may explain hypersensitivity in some autistics. Thalamic input may also be amplified if thalamic terminal fields remain the same size and therefore result in more minicolumns per stimulated macrocolumn. Each minicolumn’s response to thalamic input is also modulated by the activity of neighbouring columns to a greater or lesser degree, allowing for gradations of response, so a reduction in GABAergic inhibitory activity could also result in a loss of inhibition and greater amplification (see Figure 1). Stimuli ‘spill’ and amplification could result in increased incidence of seizures in autistics.

(Casanova 2006, pg 3, with permission)
An additional cost is that imposed by a reduction in neuron size. The ability of a neuron to sustain a connection over distance is related to the size of its cell body. Smaller neurons result in a metabolic bias favouring shorter connections at the expense of both longer distance and inter-hemispheral connectivity. The result is that autistic brains have a bias towards local (intra-regional) over global (inter-regional) connectivity and processing. Short intra-regional processing functions include mathematical calculations and visual processing. Cognitive functions that require inter-regional processing would be less metabolically efficient, including language, face recognition, and joint attention (Casanova - Abnormalities Of Cortical Circuitry In The Brains Of Autistic Individuals).
The total number of minicolumns in the human brain is defined during the first forty days of fetal development. Symmetrical divisions of germinal cells determine the total number of minicolumns, and a second phase of asymmetrical divisions provides for cell migration into the cortex, with successive divisions determining the number of cells in each minicolumn. This is a complex process, defined by a large number of genes interacting with the environment. The result is that a higher number of minicolumns is not something that someone can ‘acquire’ or be vulnerable to within the post-natal period. It is possible for disruptions during the narrow window of fetal development (e.g. rubella, thalidomide, tuberous sclerosis) to also result in an increase in the number of minicolumns, by influencing this process (Casanova - Abnormalities Of Cortical Circuitry In The Brains Of Autistic Individuals; Casanova 2006).
One point that is important to make is that while narrower minicolumns is a factor in autism, evidence suggests that it is not related to mental retardation. In Down Syndrome, as an example, minicolumn width is normal, despite the smaller brain sizes of Down Syndrome patients.
Implications
Reduced minicolumn width appears to be a prerequisite for autism. But, the reported minicolumn widths found within autistic brains are still within the normal distribution of minicolumnar width, albeit at the tail end (Casanova 2006). This suggests to me that the existence of narrow minicolumns is not enough by itself to result in an ASD diagnosis. The key instead appears to be a reduction in inhibition within minicolumns, rather than width alone. Reduced width increases the consequences of reduced inhibition, but does not automatically cause it. In effect, a brain with narrower minicolumns may be less robust, and therefore more vulnerable to the complications that could come with deviation from the narrow tolerances within which the brain functions. In a brain with wider minicolumns, a loss of inhibition would not have as significant an impact, as minicolumnar width (and therefore distance between minicolumn information processing cores) would still exist to reduce intercolumnal spill, and thalamic projections would result in fewer minicolumns per macrocolumn to be affected.
It is a logical assumption that minicolumn development in MZ twins would be similar (same alleles, very minimal epigenetic differences during the early weeks of fetal development). But MZ twins do not always share the same ASD diagnosis. In cases in which one twin has an ASD diagnosis, the chance of the second twin also having an ASD diagnosis (not necessarily of the same severity) is 60%, and the chance of the second twin being within the broader autism phenotype is 92%. While ASD clearly has an inheritable component (the prevalence of ASD in the population is 0.6%, but within families with an ASD child, the chance of subsequent children being ASD is up to 10%), genetics alone is not sufficient to explain a diagnosis in all cases. This, combined with the fact that reduced autism minicolumn widths still fall within the normal range suggests to me that the issue in autism relates to loss of inhibition, to which autistics are more vulnerable than average, i.e. reduced minicolumnar width is a precondition for autism but a 'second hit' is still required. If that second hit were purely genetic, as distinct from epigenetic or environmental, then presumably both MZ twins should be similarly affected. This is clearly not the case a significant percentage of the time.
Support for this may come from studies of the brains of MZ twins discordant for autism. In Kates et al 2003, a comparison of MZ twins discordant for autism indicated that both the autistic and non-autistic (broader autism phenotype) twins showed no significant differences in cerebral gray matter. This suggests a structural similarity in MZ twins, and the possibility of a requirement for a ‘second hit’ to explain the discordant outcomes.
We can speculate on what that ‘second hit’ may be, and indeed the speculation as to a) whether a second hit exists and b) the causes, if any, are among the most controversial subjects of debate within the community of those linked by autism. I will look a bit more closely in this direction in a post in the near future.
What is clear from above though is that if narrow minicolumnar width is a) a precondition for ASD and b) not sufficient by itself to cause ASD, then this should have an impact on ASD research. If narrow minicolumns creates a vulnerability that wider minicolumns do not, then demonstrating lack of effect in those who are invulnerable does not demonstrate lack of effect in those who may be vulnerable. In many cases the correct population in causation analyses (e.g. epidemiological studies?) would not be the wider population, but instead the sub-section of the population who are actually vulnerable. And in looking for causes of autistic disability, we may want to look more closely at those events and/or stimuli that may reduce inhibition within the brain and/or further discourage global connectivity.
It is also a reasonable proposition is that a ‘cure’ for autism would not alter the underlying narrow minicolumnar structure of the brain. Instead, it would allow this structure to function more effectively, maintaining its strengths while reducing or eliminating the more disabling features of ASD. A cure would not change who one is, which is one of the fears of the neurodiversity community. Instead, it would allow autistic thought to function without the disabling comorbidities, serving to maintain neurodiversity and allowing it to flourish.
Update - Related post:
Autism and the Evolution of the Brain (Oct 13, 2006)
Tuesday, August 29, 2006
The Agony of Waiting for Poo
Yes, dear reader, this post is about poo (bowel movements in adult speak), or more accurately, the absence of same. And please note that the ‘No Sarcasm’ sign has been turned off for the duration of this post.
Fortunately, it has been revealed by some in the neurodiversity camp that there are no proven links between any comorbidities and the causes of autism (other than - potentially - seizures). As such, I can therefore hold my head high and treat the Bear’s medical issues without any fear that I could be accused of trying to ameliorate her autism. Her biggest set of remaining medical issues - yes, there are others that now appear to have been ameliorated - are GI related, including alternating and sometimes concurrent chronic constipation and diarrhea, and absorption problems (e.g. low levels of many essential trace minerals and iron – as confirmed by Ontario government medical lab tests, for those whose thoughts veer off in rebuttal in the direction of quack labs - despite supplementation).
The latest bout (note that 'latest' is not synonymous with 'first') started on Friday at around 2:00 PM. The Bear, who had not had a bowel movement in three days, had what we affectionately call a 'shart', i.e. a small leak of gas and liquid stool. Momma Bear cleaned her up and waited for ‘the big one’ which usually follows. Instead, the Bear had another shart. Same cleanup, same wait. And another. Same cleanup, same wait. The Bear usually has a bath after a bowel movement (like night usually follows day - to us it is not a BM but a BM&B) and is fine. This time she did not, and the combination of wipes, rubbing, and irritation from the sharts left her rectum sore and red. Now comes the really fun part.
The Bear began to try harder and harder to have a bowel movement. It obviously hurt, and she started to cry. She strained harder, and cried more. And then she relaxed, calmed down, and looked just plain uncomfortable. A half hour later, the same thing, but with louder crying, screaming, and more tears. Crying is unfortunately ‘normal’ on these occasions, but this time it was worse, and the crying and straining continued on and off with increasing frequency for a few hours. Eventually she passed a small brick (yes, it was about that hard), with much crying, obvious pain, and discomfort. Momma Bear had a very hard time cleaning her up, because by this point the Bear was quite sore and resisting any and all efforts. She then had her bath, I came home from post-work grocery shopping, and we thought this was over for the evening.
But it wasn’t. Within a half an hour, she started sharting again. And pushing, and straining, and crying, and yelling. We kept checking, and trying to clean and wipe her and load on the protective barrier. Because she was so sore she kept resisting. Obviously more was coming, and the frequency of sharts was too much to put her in the bath. Faced with the choice of leaving her and allowing her to become more inflamed, or wrestling with her to clean her, we chose the second option. For some reason, the fact that we kept explaining what we were doing and saying that we were trying to help her didn’t seem to matter to her very much. And Wow! - residual hypotonia notwithstanding, is she ever strong! The frequency of attempts increased to about every ten minutes and went on for hours. We talked about taking her to the ER to see if we could get an enema (my choice), but figured that she would probably sit in the waiting room for a few hours with the same issue and no easy way for us to keep her clean, which would lead to further irritation (Momma Bear’s thoughts).
By about 3:00 AM she fell asleep, exhausted. For about ten minutes. Then she woke up, crying and whimpering as she strained, and fell back asleep. This continued all night. By about 8:00 AM, when the Bear woke up for good, Momma Bear and I were at wits end. When we checked her diaper, sure enough she had had more sharts during the night. More cleanup, more wrestling. Of course, we were handling this like the mature (and well rested) adults that we are (if you believe this…), which was helping the situation immensely. We compromised on phoning the provincial Telehealth service, where we could speak to an RN.
The RN walked us through the emergency list (still breathing - from constipation??? - and no fever, which was good), and ensured the Bear was properly hydrated (she was). She asked about her diet, which contained all the right foods, etc., and we mentioned that a) the diet was worked out by a PhD in Nutrition and b) that the Bear was in the process of coming under the care of a GI specialist at a local children’s hospital (thankfully, as a corollary of free medical care everything works at lighting speed). She suggested monitoring the situation (our first choice too for a distressed child who is regularly screaming out in pain) and bringing her to the Doctor if it didn’t clear up by the next day. She gave us a list of preferred foods (pretty much all of which the Bear already eats). She also asked if we had tried a suppository. "Er, no?" Given my experience with such things, she explained how to use one – “You want me to do what?” – and off I went to the Pharmacist to buy some.
Back home, we went through another cycle of pushing, and then we cleaned the Bear up and tried the suppository. (Of note, this is the first ‘supplement’ that I’ve tried on her that I haven’t tried on myself first). Of course, the Bear graciously cooperated through this. We put a diaper on and waited. Within five minutes she gave a mighty strain, complete with screaming and tears, and there it was – ‘Brick, the Sequel’, complete with an embedded and undissolved suppository. We were overjoyed. By early afternoon the Bear was cleaned up again, bathed, fed, and she was happy. Much happier than anyone with no real sleep the night before had any right to be. She ran around giggling, wanted to play, wrestle, was in great spirits, and still actually liked us (I’m still expecting PTSD to kick in within a few days). Now all we had to worry about was the two jars of prunes and the spoonfuls of Lansoyl. We didn’t want any more action for at least 24 hours so that the cream and time could have their healing effect.
Sunday morning, and all was still quiet. Then the strains and crying started again. And the pushing. And the sharts. She also started toe-walking, something that she had stopped doing nearly a year ago. Here we go again. This time though, we had the suppositories. We waited about half an hour to see if the Bear could resolve the problem. Momma Bear wanted to wait a few minutes more, but not this time. In went the suppository (again, with the Bear’s complete and total cooperation), and within five minutes, out came Brick #3. After the cleanup and bath, peace and order was restored. For now.
So far, everything is okay. The GI specialist has moved much higher up our priority list, and we’re trying to figure out just how many prunes one little girl can eat. But thankfully I can rest assured that – despite a lack of research (only now is the UC at Davis starting to look seriously and systematically at the issue of comorbidities) – like all of the Bear’s many medical conditions that we have cleared up (commensurate – totally coincidentally - with some of her most significant gains), this has nothing whatsoever to do with her autism. What a relief!
The ‘No Sarcasm’ sign has now been turned back on.
---
Update – Yesterday the prunes and Lansoyl caught up. The Bear had three 'normal' (for her) mushy wet BM&Bs, without a lot of upset. We’re now back to the other end of the BM spectrum, until the pendulum swings back to constipation. Note also that I promise to do my best not to write about this issue again.
Fortunately, it has been revealed by some in the neurodiversity camp that there are no proven links between any comorbidities and the causes of autism (other than - potentially - seizures). As such, I can therefore hold my head high and treat the Bear’s medical issues without any fear that I could be accused of trying to ameliorate her autism. Her biggest set of remaining medical issues - yes, there are others that now appear to have been ameliorated - are GI related, including alternating and sometimes concurrent chronic constipation and diarrhea, and absorption problems (e.g. low levels of many essential trace minerals and iron – as confirmed by Ontario government medical lab tests, for those whose thoughts veer off in rebuttal in the direction of quack labs - despite supplementation).
The latest bout (note that 'latest' is not synonymous with 'first') started on Friday at around 2:00 PM. The Bear, who had not had a bowel movement in three days, had what we affectionately call a 'shart', i.e. a small leak of gas and liquid stool. Momma Bear cleaned her up and waited for ‘the big one’ which usually follows. Instead, the Bear had another shart. Same cleanup, same wait. And another. Same cleanup, same wait. The Bear usually has a bath after a bowel movement (like night usually follows day - to us it is not a BM but a BM&B) and is fine. This time she did not, and the combination of wipes, rubbing, and irritation from the sharts left her rectum sore and red. Now comes the really fun part.
The Bear began to try harder and harder to have a bowel movement. It obviously hurt, and she started to cry. She strained harder, and cried more. And then she relaxed, calmed down, and looked just plain uncomfortable. A half hour later, the same thing, but with louder crying, screaming, and more tears. Crying is unfortunately ‘normal’ on these occasions, but this time it was worse, and the crying and straining continued on and off with increasing frequency for a few hours. Eventually she passed a small brick (yes, it was about that hard), with much crying, obvious pain, and discomfort. Momma Bear had a very hard time cleaning her up, because by this point the Bear was quite sore and resisting any and all efforts. She then had her bath, I came home from post-work grocery shopping, and we thought this was over for the evening.
But it wasn’t. Within a half an hour, she started sharting again. And pushing, and straining, and crying, and yelling. We kept checking, and trying to clean and wipe her and load on the protective barrier. Because she was so sore she kept resisting. Obviously more was coming, and the frequency of sharts was too much to put her in the bath. Faced with the choice of leaving her and allowing her to become more inflamed, or wrestling with her to clean her, we chose the second option. For some reason, the fact that we kept explaining what we were doing and saying that we were trying to help her didn’t seem to matter to her very much. And Wow! - residual hypotonia notwithstanding, is she ever strong! The frequency of attempts increased to about every ten minutes and went on for hours. We talked about taking her to the ER to see if we could get an enema (my choice), but figured that she would probably sit in the waiting room for a few hours with the same issue and no easy way for us to keep her clean, which would lead to further irritation (Momma Bear’s thoughts).
By about 3:00 AM she fell asleep, exhausted. For about ten minutes. Then she woke up, crying and whimpering as she strained, and fell back asleep. This continued all night. By about 8:00 AM, when the Bear woke up for good, Momma Bear and I were at wits end. When we checked her diaper, sure enough she had had more sharts during the night. More cleanup, more wrestling. Of course, we were handling this like the mature (and well rested) adults that we are (if you believe this…), which was helping the situation immensely. We compromised on phoning the provincial Telehealth service, where we could speak to an RN.
The RN walked us through the emergency list (still breathing - from constipation??? - and no fever, which was good), and ensured the Bear was properly hydrated (she was). She asked about her diet, which contained all the right foods, etc., and we mentioned that a) the diet was worked out by a PhD in Nutrition and b) that the Bear was in the process of coming under the care of a GI specialist at a local children’s hospital (thankfully, as a corollary of free medical care everything works at lighting speed). She suggested monitoring the situation (our first choice too for a distressed child who is regularly screaming out in pain) and bringing her to the Doctor if it didn’t clear up by the next day. She gave us a list of preferred foods (pretty much all of which the Bear already eats). She also asked if we had tried a suppository. "Er, no?" Given my experience with such things, she explained how to use one – “You want me to do what?” – and off I went to the Pharmacist to buy some.
Back home, we went through another cycle of pushing, and then we cleaned the Bear up and tried the suppository. (Of note, this is the first ‘supplement’ that I’ve tried on her that I haven’t tried on myself first). Of course, the Bear graciously cooperated through this. We put a diaper on and waited. Within five minutes she gave a mighty strain, complete with screaming and tears, and there it was – ‘Brick, the Sequel’, complete with an embedded and undissolved suppository. We were overjoyed. By early afternoon the Bear was cleaned up again, bathed, fed, and she was happy. Much happier than anyone with no real sleep the night before had any right to be. She ran around giggling, wanted to play, wrestle, was in great spirits, and still actually liked us (I’m still expecting PTSD to kick in within a few days). Now all we had to worry about was the two jars of prunes and the spoonfuls of Lansoyl. We didn’t want any more action for at least 24 hours so that the cream and time could have their healing effect.
Sunday morning, and all was still quiet. Then the strains and crying started again. And the pushing. And the sharts. She also started toe-walking, something that she had stopped doing nearly a year ago. Here we go again. This time though, we had the suppositories. We waited about half an hour to see if the Bear could resolve the problem. Momma Bear wanted to wait a few minutes more, but not this time. In went the suppository (again, with the Bear’s complete and total cooperation), and within five minutes, out came Brick #3. After the cleanup and bath, peace and order was restored. For now.
So far, everything is okay. The GI specialist has moved much higher up our priority list, and we’re trying to figure out just how many prunes one little girl can eat. But thankfully I can rest assured that – despite a lack of research (only now is the UC at Davis starting to look seriously and systematically at the issue of comorbidities) – like all of the Bear’s many medical conditions that we have cleared up (commensurate – totally coincidentally - with some of her most significant gains), this has nothing whatsoever to do with her autism. What a relief!
The ‘No Sarcasm’ sign has now been turned back on.
---
Update – Yesterday the prunes and Lansoyl caught up. The Bear had three 'normal' (for her) mushy wet BM&Bs, without a lot of upset. We’re now back to the other end of the BM spectrum, until the pendulum swings back to constipation. Note also that I promise to do my best not to write about this issue again.
Friday, August 18, 2006
A Couple of Bear Updates
Sometimes you lose, and sometimes you win.
On Wednesday the Bear had another PECS session. She was doing so well at home, demonstrating that she understood which card to use to request ‘milk’, or her soother, or the swing, and she was also showing signs that she was starting to understand other cards as well (for various videos and games we play with her).
So at the PECS session, I set up her binder with three cards on it (nose, spoon, and ‘milk’). She selected the milk card and handed it to me. I held it up to show her, said ‘milk’, and then handed her the cup with a couple of ounces of milk in it. While she drank the 'milk' I put the binder out again with the soother card. When she finished, she handed me the cup, looked over, grabbed the soother card, and handed it to me. I again showed her the card, said ‘soother’ and gave it to her. So far so good.
The therapist then said, what would happen if you gave her a choice of milk or soother, and then had her choose the right item after giving me the card? I was confident that she would get this right. We put both cards on the binder, and she pulled the milk card. I then showed her the card and identified it, and put both options in front of her. She scooped up the soother and walked away.
We repeated this three times, and each time she selected the milk card and then took the soother. Apparently ‘milk’ is the first card to choose in any situation, followed by soother, and then by something else.
We’ll keep working on this one. At least she can distinguish between the cards. Now we just have to link them to the correct object/action.
---
When the Bear goes to IBI we always pack her a snack. Today was sliced organic strawberries and kiwi. The goal is to get her to use a pincer grasp to pick up the fruit and eat it. If they put the food in her hand she will put it in her mouth, but getting her to pick it up herself is not easy, and a pincer grasp is harder still. Most of the time she will grab the tutor’s hand and guide it to the bowl to signal that she wants them to pick up the food for her and put it in her hand. We and they also put food on her spoon and try to get her (guided) to put it in her mouth. She has never attempted to do this by herself.
Today, totally unprompted, she reached over by herself and picked up the spoon, which had a piece of fruit on it, and put it in her mouth, as if she had been doing it for years. Her school is closed for vacation for the next two weeks. In the meantime, you can bet that she will have lots of access to a bowl of sliced fruit and a spoon.
Sometimes you get a win, and sometimes you don’t. I’ll put this week in the win column. Tonight we’re off to Canada’s Wonderland to celebrate.
On Wednesday the Bear had another PECS session. She was doing so well at home, demonstrating that she understood which card to use to request ‘milk’, or her soother, or the swing, and she was also showing signs that she was starting to understand other cards as well (for various videos and games we play with her).
So at the PECS session, I set up her binder with three cards on it (nose, spoon, and ‘milk’). She selected the milk card and handed it to me. I held it up to show her, said ‘milk’, and then handed her the cup with a couple of ounces of milk in it. While she drank the 'milk' I put the binder out again with the soother card. When she finished, she handed me the cup, looked over, grabbed the soother card, and handed it to me. I again showed her the card, said ‘soother’ and gave it to her. So far so good.
The therapist then said, what would happen if you gave her a choice of milk or soother, and then had her choose the right item after giving me the card? I was confident that she would get this right. We put both cards on the binder, and she pulled the milk card. I then showed her the card and identified it, and put both options in front of her. She scooped up the soother and walked away.
We repeated this three times, and each time she selected the milk card and then took the soother. Apparently ‘milk’ is the first card to choose in any situation, followed by soother, and then by something else.
We’ll keep working on this one. At least she can distinguish between the cards. Now we just have to link them to the correct object/action.
---
When the Bear goes to IBI we always pack her a snack. Today was sliced organic strawberries and kiwi. The goal is to get her to use a pincer grasp to pick up the fruit and eat it. If they put the food in her hand she will put it in her mouth, but getting her to pick it up herself is not easy, and a pincer grasp is harder still. Most of the time she will grab the tutor’s hand and guide it to the bowl to signal that she wants them to pick up the food for her and put it in her hand. We and they also put food on her spoon and try to get her (guided) to put it in her mouth. She has never attempted to do this by herself.
Today, totally unprompted, she reached over by herself and picked up the spoon, which had a piece of fruit on it, and put it in her mouth, as if she had been doing it for years. Her school is closed for vacation for the next two weeks. In the meantime, you can bet that she will have lots of access to a bowl of sliced fruit and a spoon.
Sometimes you get a win, and sometimes you don’t. I’ll put this week in the win column. Tonight we’re off to Canada’s Wonderland to celebrate.
Saturday, August 12, 2006
Five Weird Things
As part of the meme going around the bloggosphere, Wade ‘tagged’ me to identify five weird things about myself. Narrowing down the list was hard – e.g. do the giant one-eyed white cat (now deceased), and the constant presence of Lithuanians in my life count? Where is Lithuania, anyway? (Don’t worry, I know exactly where it is!) But here goes:
1. I can’t swim. Yes, I know Canada is cold at least one month of the year, but it does have a lot of lakes, ocean shoreline, swimming pools, etc., plus the colony of Florida, so the opportunity is there. Despite repeated attempts to learn, I cannot master the breathing technique involved. I love the water, I love to canoe (and am quite fast), I’ve done more than a little portaging and whitewater, and can finally make my way across the length of a small backyard pool. But that is my limit. Fortunately, Momma Bear has every lifeguard designation imaginable. The Bear loves the water, so we’ll see who’s genes she has inherited.
2. I like to travel on my own. While I also enjoy vacations with others, and have gone away with family, large groups, small groups, girlfriends, and now Momma Bear (not yet with the Bear, but soon), I’m quite comfortable going on my own. I went to Israel by myself and worked on a kibbutz for four months (not a southern kibbutz where one hangs out with other non-Israelis, but a smaller northern kibbutz where I was at times the only volunteer). I’ve also backpacked and EurRailed around Europe by myself, from Norway and Finland to Greece and Italy, including through what was once East Germany and Yugoslavia. I also studied abroad with a girlfriend, and when we broke up I again went off traveling by myself for a bit. Whenever I wanted company it was never an issue to meet people and temporarily travel with them, but when it was time to move on, so be it. I would feel quite comfortable doing the same today, but confess that I would much rather go with the Bear and Momma Bear.
3. I love cartoons. I’ll date myself by mentioning The Flintstones, The Jetsons, Tom & Jerry, Bugs Bunny (for which I will drop almost anything), Tweety & Sylvester, Yosemite Sam, the Roadrunner & Wile E Coyote, etc. I’m not so big on the more up-to-date stuff, but I will confess to having to be dragged out of Entertainment departments in stores when 'Ice Age' or 'Monsters, Inc' have been playing.
4. I designed our house. I don’t mean rough sketches, but a proper blueprint-ready design. When we were ready to trade up, my wife and I hunted on weekends for a house that we liked. She also went looking during the week. For two years she scoured our region (we had the gas bills and mileage to prove it). When we couldn’t find anything, she switched to looking for land so that we could build to our own design. Once we found land, we went looking for a design, but again couldn’t find anything that was ‘just right’, so I got a couple of good books on design, figured out how to do it (including sizing – e.g. size rooms to match construction material sizing to keep costs in line), and drew up my own design (obviously with input and approval from Momma Bear). When done, we found a contractor to build it, and he turned it into final blueprints first. He didn’t have to change a thing.
My only regret is that the master bedroom is downstairs, while the rest of the bedrooms are upstairs. The theory was that the kids would be five and under for five years and teenagers for seven. I’m paying for that now.
5. I won a weight loss contest, losing 46 lbs in 77 days. We had a contest at work to see who could lose the largest percentage of their starting body weight. The contest ran from early January to end of March. Everyone put in $100 each, and the person who won took the pot, with second and third place getting their money back. There were twelve of us. I weighed in at 197 lbs (I’m 6 feet tall). It was during the time that we were building the house, and I wanted to use the money to pay for a Jaccuzi. I’m also way too cheap to give away $100. So I dieted (proper nutrition in small quantities and vitamins), exercised (at one point I was walking up 100 floors per day – yes, 100 floors), and the weight melted away. I had no idea how the others were doing except during the two early weigh-ins (in which a couple of others were close), so I just kept going. By the end of the contest I was down to 151 lbs, which in perspective is four pounds less than I weighed in undergrad. No one else was even close. Yes, I can be a bit competitive.
I used the money to buy a two person Jaccuzi (well, the contest paid for most of it). We’ve used the Jacuzzi three times in five years (all in the first two months). Today it holds the Bear’s bath tub, where my wife baths her. I now weigh around 180 lbs, having put most of the weight back on within three months.
So, now it’s my turn to tag three people. I tag:
María Luján (I’ll guest host if you want, but I see no reason that you should escape this)
Kristina
Shawn
Sorry all.
And Wade, starting a new meme tradition, in retaliation I just signed you up for the Lithuanian Wine of the Month club. ;-)
1. I can’t swim. Yes, I know Canada is cold at least one month of the year, but it does have a lot of lakes, ocean shoreline, swimming pools, etc., plus the colony of Florida, so the opportunity is there. Despite repeated attempts to learn, I cannot master the breathing technique involved. I love the water, I love to canoe (and am quite fast), I’ve done more than a little portaging and whitewater, and can finally make my way across the length of a small backyard pool. But that is my limit. Fortunately, Momma Bear has every lifeguard designation imaginable. The Bear loves the water, so we’ll see who’s genes she has inherited.
2. I like to travel on my own. While I also enjoy vacations with others, and have gone away with family, large groups, small groups, girlfriends, and now Momma Bear (not yet with the Bear, but soon), I’m quite comfortable going on my own. I went to Israel by myself and worked on a kibbutz for four months (not a southern kibbutz where one hangs out with other non-Israelis, but a smaller northern kibbutz where I was at times the only volunteer). I’ve also backpacked and EurRailed around Europe by myself, from Norway and Finland to Greece and Italy, including through what was once East Germany and Yugoslavia. I also studied abroad with a girlfriend, and when we broke up I again went off traveling by myself for a bit. Whenever I wanted company it was never an issue to meet people and temporarily travel with them, but when it was time to move on, so be it. I would feel quite comfortable doing the same today, but confess that I would much rather go with the Bear and Momma Bear.
3. I love cartoons. I’ll date myself by mentioning The Flintstones, The Jetsons, Tom & Jerry, Bugs Bunny (for which I will drop almost anything), Tweety & Sylvester, Yosemite Sam, the Roadrunner & Wile E Coyote, etc. I’m not so big on the more up-to-date stuff, but I will confess to having to be dragged out of Entertainment departments in stores when 'Ice Age' or 'Monsters, Inc' have been playing.
4. I designed our house. I don’t mean rough sketches, but a proper blueprint-ready design. When we were ready to trade up, my wife and I hunted on weekends for a house that we liked. She also went looking during the week. For two years she scoured our region (we had the gas bills and mileage to prove it). When we couldn’t find anything, she switched to looking for land so that we could build to our own design. Once we found land, we went looking for a design, but again couldn’t find anything that was ‘just right’, so I got a couple of good books on design, figured out how to do it (including sizing – e.g. size rooms to match construction material sizing to keep costs in line), and drew up my own design (obviously with input and approval from Momma Bear). When done, we found a contractor to build it, and he turned it into final blueprints first. He didn’t have to change a thing.
My only regret is that the master bedroom is downstairs, while the rest of the bedrooms are upstairs. The theory was that the kids would be five and under for five years and teenagers for seven. I’m paying for that now.
5. I won a weight loss contest, losing 46 lbs in 77 days. We had a contest at work to see who could lose the largest percentage of their starting body weight. The contest ran from early January to end of March. Everyone put in $100 each, and the person who won took the pot, with second and third place getting their money back. There were twelve of us. I weighed in at 197 lbs (I’m 6 feet tall). It was during the time that we were building the house, and I wanted to use the money to pay for a Jaccuzi. I’m also way too cheap to give away $100. So I dieted (proper nutrition in small quantities and vitamins), exercised (at one point I was walking up 100 floors per day – yes, 100 floors), and the weight melted away. I had no idea how the others were doing except during the two early weigh-ins (in which a couple of others were close), so I just kept going. By the end of the contest I was down to 151 lbs, which in perspective is four pounds less than I weighed in undergrad. No one else was even close. Yes, I can be a bit competitive.
I used the money to buy a two person Jaccuzi (well, the contest paid for most of it). We’ve used the Jacuzzi three times in five years (all in the first two months). Today it holds the Bear’s bath tub, where my wife baths her. I now weigh around 180 lbs, having put most of the weight back on within three months.
So, now it’s my turn to tag three people. I tag:
María Luján (I’ll guest host if you want, but I see no reason that you should escape this)
Kristina
Shawn
Sorry all.
And Wade, starting a new meme tradition, in retaliation I just signed you up for the Lithuanian Wine of the Month club. ;-)
Subscribe to:
Posts (Atom)
